Informativo Técnico Nº. 165
Treonina digestível em rações para matrizes
suínas em lactação.
Autor : Vladimir Fortes de Oliveira; Juarez Donzele; Márvio Lobão Teixeira de
Abreu; Rita Flávia Miranda de Oliveira; Bruno Alexander Nunes Silva; Alysson Saraiva.
Sumário
RESUMO
Sessenta matrizes suínas pluríparas
(média de quatro partos) de linhagem comercial, com peso inicial de 262,7 ±
Palavras-chave: aminoácidos,
exigências nutricionais, suínos
Introdução
O baixo consumo de ração pelas fêmeas suínas na
fase de lactação pode ocasionar perda de peso e prejudicar os
desempenhos produtivo e reprodutivo da matriz. A perda de peso de
matrizes lactantes é composta principalmente de tecido adiposo e protéico e
pode levar a maior intervalo desmama-cio, menor taxa
de parto e menor número de leitões
nascidos, em razão da menor taxa de ovulação (Andries,
2004).
Estudando o impacto sobre a nutrição de
aminoácidos durante a lactação, Jones & Stahly
(1999) concluíram que a restrição de aminoácidos durante a lactação aumenta a
mobilização de tecido protéico de matrizes suínas.
A perda excessiva de proteína é responsável por
metade dos prejuízos na reprodução pós-desmama, enquanto a perda excessiva de
gordura corporal representa menos de um quarto dos danos reprodutivos nesse
mesmo período (Clowes et al., 2005).
Além do consumo de aminoácidos, o balanço aminoacídico das rações também tem sido estudado. Segundo
Kim et al. (2001), em
matrizes com baixo consumo de ração e alta taxa de mobilização tecFidual durante a lactação, a treonina
é o segundo aminoácido limitante. Por outro lado, em matrizes com alto consumo
durante a lactação e baixa taxa de mobilização, a valina é o segundo aminoácido
limitante em rações à base de milho e farelo de soja. Em ambos os casos, a
lisina é o primeiro aminoácido limitante. Desse modo, a ordem dos aminoácidos
limitantes está relacionada ao que se pode esperar em mobilização de tecidos
das matrizes durante a lactação.
No Brasil, as médias de temperatura são mais
altas que as de países de clima temperado, o que limita o consumo de ração
pelas matrizes suínas, principalmente no verão. Assim, são observados consumos de
ração próximos de 5 kg/dia, valor inferior ao de 7 kg/dia observado em países
de clima temperado.
Este estudo foi conduzido para avaliar níveis de treonina digestível em rações para matrizes em lactação,
considerando os desempenhos produtivo e reprodutivo da matriz
e o desempenho da leitegada.
Material e Métodos
O experimento foi conduzido na
Fazenda Água Limpa, localizada em Jequeri, Minas
Gerais, em julho de 2004. Utilizaram-se 60 matrizes suínas híbridas de
linhagem comercial (Landrace × Large
White) distribuídas em delineamento experimental inteiramente casualizado com 4 tratamentos e 15 repetições, considerando
a matriz a unidade experimental. Como tratamentos, avaliaram-se rações com 0,608; 0,646; 0,684 ou 0,722% de treonina
digestível, correspondentes às relações treonina
digestível:lisina digestível de 64, 68, 72 e 76%, respectivamente.
As matrizes foram distribuídas nos tratamentos de
acordo com o peso e a ordem de parto, de forma a se obter maior homogeneidade
entre os tratamentos. Para isso, foram agrupadas segundo a ordem de parto: no
primeiro grupo, foram incluídas as fêmeas de primeiro parto; no segundo, as
fêmeas de segundo a sétimo partos; e no terceiro, as
matrizes de sétimo parto em diante. As matrizes de cada grupo foram
distribuídas, eqüitativamente, nos níveis de treonina
de forma que em cada grupo fosse mantido o mesmo número de fêmeas de cada um
dos três grupos por ordem de parto.
As rações experimentais foram formuladas à base
de milho e farelo de soja e suplementadas com minerais e vitaminas (Tabela 1)
para atender às exigências nutricionais mínimas descritas por Rostagno et
al. (2000), exceto de valina e treonina.
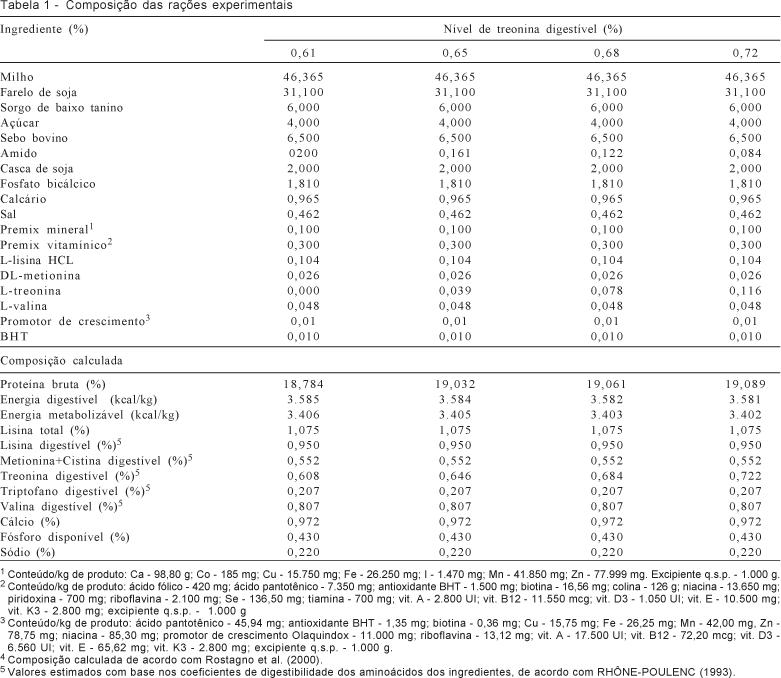
Em todas as rações, foram conferidas as relações aminoacídicas entre os demais aminoácidos essenciais
digestíveis e a lisina digestível a fim de se assegurar que, em todos os
tratamentos, nenhum outro aminoácido estivesse limitante na ração. Foram
utilizadas relações superiores às preconizadas com base na proteína ideal para
fêmeas em lactação e os níveis de treonina digestível
foram obtidos por meio da suplementação de L-treonina
(98% de treonina) em substituição proporcional ao
amido. A ração e a água foram fornecidas à vontade.
As matrizes foram pesadas no dia do parto e à desmama, que ocorreu no 16º dia após o parto. A espessura
de toucinho foi medida por meio de aparelho de ultra-som modelo RENCO Sono Grader, no dia no parto e à
desmama, a
Após a desmama, as
matrizes foram levadas ao setor de gestação, onde foi iniciado o manejo de detecção de cio. A identificação
do cio foi realizada com a presença de um macho à frente das fêmeas, que foram
testadas duas vezes por dia (manhã e tarde) quanto ao reflexo de tolerância ao
macho. O cio foi confirmado quando a fêmea se manteve estática e receptiva à
presença do macho.
Como o número de leitões pode influenciar a
produção de leite e a mobilização de proteína corporal da matriz (Kim et al., 2001), as leitegadas foram
equalizadas (10 e 11) no terceiro dia após o parto para evitar variações nas
respostas dos animais que não fossem decorrentes dos níveis de treonina estudados. A pesagem dos leitões foi realizada
após a equalização e à desmama. Os leitões foram
submetidos ao manejo tradicional (corte de dentes e cauda, cura do umbigo ao
nascimento e aplicação de ferro dextrano no terceiro
dia) e receberam a ração pré-inicial a partir do terceiro dia de vida.
Nos leitões, avaliou-se o ganho
de peso médio diário e, nas matrizes, avaliaram-se os consumos de ração, de treonina digestível e de lisina digestível, a perda de
peso, as perdas de proteína e de gordura corporal, a variação na espessura de
toucinho, a produção de leite e o intervalo desmama-estro.
A perda de proteína foi estimada considerando as
quantidades de proteína corporal no início e ao final do experimento, segundo
equação preconizada por Whittemore & Yang (1989):
proteína corporal = -2,3 + [0,19 × peso corporal (kg)] [0,22 × ETP2 (mm)].
A perda de gordura corporal foi estimada a partir das quantidades de gordura
corporal no início e ao final do experimento, segundo a equação de Whittemore & Yang (1989): gordura corporal = -20,4 +
[0,21 × peso corporal (kg)] + [1,5 × ETP2 (mm)].
A produção de leite pela porca foi estimada
utilizando-se a equação proposta por Noblet & Etienne (1989): produção de leite (g/dia) = (2,42 × ganho
de peso da leitegada (g/dia) + (78,2 × peso da leitegada à equalização (kg) +
(26 × número de leitões à desmama).
Para todos os parâmetros avaliados (matrizes e
leitões), foram considerados como períodos inicial e final do
experimento os dias da equalização dos leitões nas leitegadas e do
desmame, respectivamente.
As análises estatísticas dos dados de desempenho
produtivo e reprodutivo das matrizes e do desempenho dos leitões foram
realizadas utilizando-se o programa computacional SAEG (Sistema de Análises Estatísticas
e Genéticas), versão 8.0 (UFV, 2000), por meio dos procedimentos para análises
de variância e regressão.
Resultados e Discussão
O consumo diário de ração (CDR) das matrizes não
foi influenciado (P>0,05) pelos níveis de treonina
digestível da ração (Tabela 2). Resultados semelhantes foram observados por
Cooper et al. (2001), que,
avaliando diferentes relações (
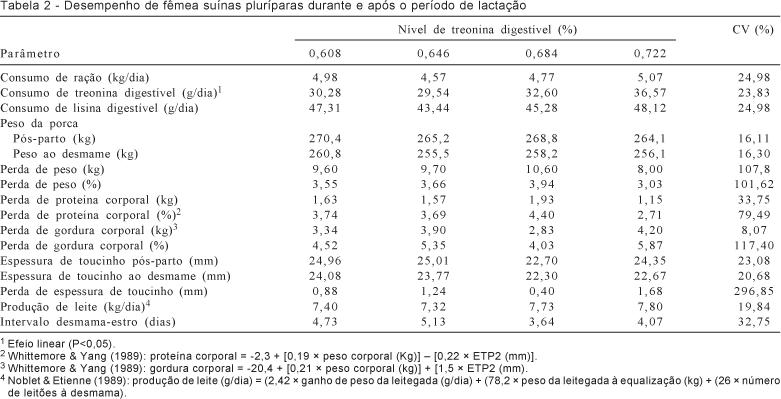
Os animais utilizados neste estudo apresentaram
consumo de ração médio diário de
O consumo de treonina
digestível aumentou de forma linear (P<0,05), segundo a equação =
-5,8364 +57,2619 X (r2 = 0,80), de
acordo com o aumento dos níveis de treonina, fato
relacionado à ausência de variação no consumo diário de ração e ao aumento dos
níveis de treonina digestível das rações. O menor
consumo de treonina digestível observado neste
estudo, de 29,54 g/dia, foi similar aos valores propostos por Cooper et al. (2001), de 29, 30 e 31
g/dia para redução da mobilização de tecido corporal de fêmeas de primeiro,
segundo e terceiro partos, respectivamente, durante a lactação.
Os níveis de treoninada
ração não influenciaram (P>0,05) o consumo de
lisina digestível pelos
animais, fato justificado pela ausência de variação
do consumo médio diário de ração
pelos animais e pelo fato de as rações terem sido isolisínicas.
As matrizes apresentaram consumo de lisina digestível médio diário de
Do mesmo, os níveis de treonina
digestível da ração não influenciaram (P>0,05) a perda de peso das matrizes
no período de lactação. Resultados semelhantes foram observados por Cooper et al. (2001), que não verificaram
variação na perda de peso corporal das matrizes quando os níveis de treonina digestível da ração variaram de
A ausência de efeito sobre a perda de peso das
porcas pode ser justificada pela falta da necessidade de mobilização de tecidos
corporais para suprir a demanda de aminoácidos para produção de leite. Segundo
Kim & Easter (2003), quando não há
disponibilidade suficiente de aminoácidos provenientes da ração, a fêmea
mobiliza tecido corporal para disponibilizar aminoácidos para produção de
leite.
A reduzida perda de peso dos animais neste estudo
pode ser explicada pela duração do período de lactação estudado e pelo perfil
nutricional das rações experimentais, mais precisamente seu conteúdo em lisina.
Estudos têm comprovado (Cooper et al., 2001) que o período de
maior desgaste para a fêmea é a terceira semana de lactação, quando a demanda
por nutrientes para a produção de leite é mais alta. Como as porcas neste
experimento foram desmamadas em média aos 16 dias, é possível que os animais
não tenham experimentado este período de maior desgaste corporal.
O consumo de lisina pelas porcas em lactação é um
parâmetro importante na determinação do grau de mobilização de tecido corporal.
Segundo Clowes et
al. (2003),
Em condições de intensa mobilização de tecido
corporal, a treonina passa a ser o aminoácido mais
limitante, depois da lisina, em rações para porcas em lactação (Kim et al., 2001; Kevin et al., 2003; Soltwedel et al., 2003). Estudando relações entre treonina
digestível e lisina digestível, Kim et
al. (2001) observaram que, para matrizes cuja perda de peso não é significativa
(abaixo de 12% do peso corporal), a relação entre treonina
digestível: lisina digestível é de 59%, enquanto, para fêmeas com perdas de
peso de 30%, a relação entre esses aminoácidos é de 62%.
As perdas de proteína corporal, tanto absoluta
como relativa, não foram influenciadas (P>0,05) pelos níveis de treonina digestível estudados, o que está coerente com os
resultados de perda de peso corporal dos animais. Uma vez que as necessidades
de aminoácidos dos animais para mantença e produção de leite foram atendidas
quase integralmente pela dieta, a mobilização de proteína corporal durante todo
o período estudado foi em média de
Como observado para perda de proteína corporal,
os níveis de treonina digestível da ração também não
influenciaram (P>0,05) a perda de gordura corporal dos animais, resultado
justificado também pela ausência de efeito dos níveis de treonina
da dieta sobre a perda de peso corporal das matrizes durante o período
estudado. Do mesmo modo, Johnston et
al. (1993) analisaram a resposta de matrizes suínas aos níveis protéicos
durante a lactação e verificaram que não houve diferença significativa na perda
de gordura corporal desses animais.
A espessura de toucinho pós-desmame e a perda de
espessura de toucinho dos animais não foram afetadas (P>0,05) pelos níveis
de treonina da ração, o que está de acordo com os
dados observados para perda de gordura corporal. Resultados semelhantes foram
obtidos por Cooper et al.
(2001), que, avaliando níveis de treonina para
animais de alta capacidade produtiva em fase de lactação, não observaram
diferença significativa na espessura de toucinho no final do período de
lactação.
Não se observou efeito (P>0,05) dos níveis de treonina sobre a produção de leite das matrizes. Essa
produção, afetada principalmente pelo consumo de energia, é pouco sensível aos
níveis de aminoácidos ingeridos, uma vez que a mobilização de proteína corporal
pode atender a necessidade de aminoácidos para a síntese de leite. A ausência de diferenças no consumo de ração das porcas entre os
níveis de treonina avaliados e o fato de as rações
serem isocalóricas justificam a ausência de
efeito dos níveis de treonina da ração sobre a
produção de leite dos animais.
O intervalo desmama-estro dos animais também não
foi influenciado pelos níveis de treonina digestível
das rações, o que confirma relatos de Clowes et al. (2003) de que a matriz
suína lactante é capaz de suportar até 12% de perda de peso corporal (a perda
observada neste estudo foi em média de 3,6%) sem prejuízos na função
reprodutiva.
O prejuízo na função
reprodutiva pode ser explicado, em parte, pelo fato de que, durante a
mobilização de proteína corporal para atendimento das necessidades de
aminoácidos da fêmea, a maior taxa de mobilização de proteína tecidual (em
porcentagem de peso) ocorre no trato reprodutivo e o tecido muscular é o maior
doador quantitativo de aminoácidos (Kim & Easter,
2003). Ainda segundo Clowes et al. (2003), altas taxas de
perda de proteína corporal estão associadas às dificuldades na maturação
nuclear e citoplasmática de oócitos in vitro, o que não parece ser o caso das matrizes em
estudo, pois o intervalo desmama-estro médio observado (de 4,39 dias) pode ser
considerado adequado para sistemas de criação intensiva de suínos, portanto, é
indicativo de que não houve prejuízo da função reprodutiva dos animais.
Durante o período de lactação, o metabolismo de
aminoácidos prioriza a produção de leite (Kim & Easter,
2003), de modo que o não atendimento das necessidades em aminoácidos por meio
da ração provoca mobilização de proteína do corpo do animal. O grau de
mobilização tecidual observado (Tabela 3) indica que a necessidade de
aminoácidos dos animais foi quase completamente atendida pela dieta. O consumo
de lisina médio diário observado, de
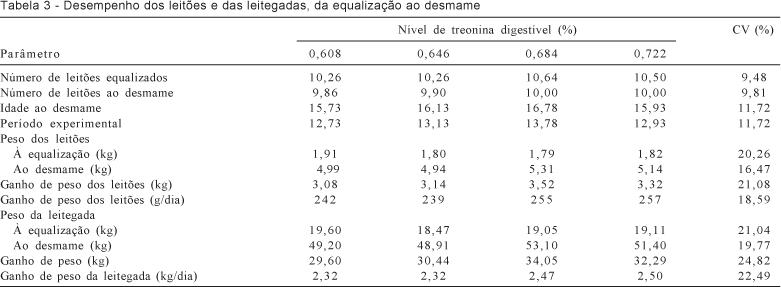
O número de leitões desmamados não variou
(P>0,05) entre os níveis de treonina digestível
estudados, o que significa que as rações não influenciaram a taxa de
mortalidade dos leitões durante a lactação.
O ganho de peso dos leitões e das leitegadas não
foi influenciado (P>0,05) pelo aumento do consumo de treonina
pelas matrizes em lactação, o que está coerente com os dados de produção de
leite das porcas neste estudo. Silva et
al. (2003) também não observaram diferenças no desempenho das leitegadas quando
aumentaram a relação treonina digestível:lisina
digestível da ração.
Conclusões
O nível de 0,608% de treonina digestível, correspondente à relação treonina digestível:lisina digestível de 64%, atende às
exigências produtivas e reprodutivas de treonina
digestível de matrizes suínas lactantes com perdas de peso de 3,6% durante
período médio de lactação de 16,14 dias.