Informativo Técnico Nº. 169
Interação entre sistema imunologico do suino e micotoxicose.
Autor : Prof. Elizabeth Santin.
Introdução
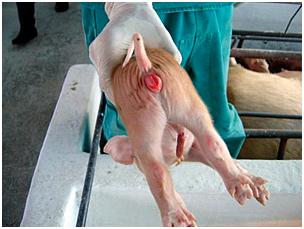
A presença de micotoxinas
na ração, podem desencadear alguns sinais clínicos
como o edema de vulva em leitões recém-nascidos.
O sistema imunológico dos animais é fundamental
para a defesa contra agentes estranhos ao organismo como vírus,
bactérias, protozoários e outros parasitas. Na realidade, são o
equilíbrio entre a boa funcio-nalidade deste sistema
(imuno-competência) e os desafios de campo que vão
determinar a saúde de um animal. Sempre que ocorrer aumento na pressão infectiva ou uma diminuição na capacidade imunogênica do
indivíduo ocorre o que caracterizamos como doença.
Animais enfermos são menos produtivos, e, além
disso, aumentam os custos de produção por necessidade de medicamentos e intervenções veterinárias.
Devido a isso, atualmente vários programas de manejo
(incluindo programas de biossegurança e criações all in all out) têm colaborado na
diminuição da pressão infectiva nas instalações para
animais. Entretanto, existem vários fatores nesta forma de produção intensiva
de animais que possibilita uma menor competência do sistema imunológico dos
animais como situações de estresse ou presença de fatores anti-nutricionais
e tóxicos na ração. Entre estes fatores destaca-se o papel das micotoxinas, que são produtos do metabolismo intermediário
de fungos e que podem estar presentes na ração. O crescimento fúngico em cereais e rações é cada vez mais comum devido as novas tecnologias de plantio de cereais, péssimas
condições de transporte, armazenamento e estocagem destas matérias primais, o
que aumenta muito a probabilidade da presença de micotoxinas
nas rações.
Essas micotoxinas
presentes na ração podem causar diversas alterações metabólicas no animal, que
acarretam em diversas apresentações de enfermidades que chamamos micotoxicoses. Entre essas micotoxicoses
clínicas específicas, podem-se citar as conhecidas formas clínicas
caracterizadas por estrogenismos e abortos provocados
por zearalenona, o edema pulmonar porcino
provocado por fumonisinas, vômitos provocados pela
toxina DON (vomitoxina), etc. O grande problema no
que se refere as micotoxinas
é que doses de micotoxinas que não causam esses
quadros clínicos conhecidos podem culminar com o aparecimento de
imunossupressão nos animais caracterizadas por infecções subclínicas,
falhas de vacinações e enfermidades infecciosa nos animais. Vários são os
mecanismos pelos quais as distintas micotoxinas
conhecidas podem atuar no sistema imunológico. Desta maneira, neste texto,
busca-se apresentar estudos envolvendo a interferência destas micotoxinas com o sistema imunológico de suínos.
Conhecendo Sistema Imunológico em Suínos
De acordo com Abbas et al.,(2000), o termo imunidade
tem sua origem na palavra latina immunitas que na
Roma antiga referia-se a exceções legais oferecidas aos senadores da época
durante seu período de mandato. Infelizmente hoje ainda esses privilégios
políticos acabam trazendo inúmeros problemas para nossa sociedade moderna.
Entretanto, a parte disso, ao longo dos séculos o termo imunidade refere-se a
prevenir doenças, mais especificamente a doenças infecciosas. Uma definição mais
ampla de imunidade relaciona a reação do organismo contra substâncias incluindo
microorganismos, macromoléculas como proteínas e polissacarídeos. As células e
moléculas responsáveis por essa imunidade constituem o sistema imunológico e a
interação coletiva dessas estruturas e moléculas na resposta contra agentes
estranhos ao organismo é denominado resposta
imunológica.

Quando o sistema imunológico funciona adequadamente
e ocorre um equilíbrio entre ele e a pressão infecciosa, o animal encontra-se
em condições de homeostase ou apresenta-se saudável.
Em condições de homeostase, este sistema é um
consumidor modesto dos recursos orgânicos, porém, uma vez ativado, torna-se um
grande sorvedouro de energia e nutrientes - os quais o organismo animal desvia
da produção de carne, leite e ovos (Koutsos e Klasing, 2001). Algumas vezes pode ocorrer também,
que esses mecanismos desenvolvidos para defender o organismo causem danos
teciduais e doenças.
A defesa do organismo contra agentes estranhos é
mediada por reações primárias da imunidade inata do organismo. Posteriormente,
ocorre a resposta imune denominada adaptativa. A imunidade inata é reconhecida
como barreiras físicas e químicas como epitélio e substâncias produzidas por
algumas células que possam ter alguma atividade anti-bacteriana
como o ácido estomacal, por exemplo.
Além disso, fazem ainda parte da imunidade inata
do organismo as células fagocíticas (macrófagos e
neutrófilos) e células natural killer (NK), proteínas do sangue como as que
compõem o sistema complemento e as citocinas que
regulam e coordenam muitas atividades das células que fazem parte desta
imunidade inata. Apesar da imunidade inata já existir no animal, independente
de qualquer desafio, sabe-se que esta imunidade aumenta em magnitude e
capacidade defensiva a cada exposição sucessiva a distintos
agentes.
Por outro lado, a imunidade
adaptativa é considerada a imunidade desenvolvida especificamente para um
agente, por isso também é denominada imunidade específica, e tem a capacidade
de “memorizar”, de forma que a segunda resposta a esse mesmo agente será ainda
mais vigorosa que da primeira vez em que esse agente for detectado. Os
componentes primordiais desta resposta específica são os linfócitos e seus
produtos como as linfocinas e os anticorpos. Na
realidade, estes dois tipos de imunidade (inata e adaptativa) trabalham de
forma integrada, sendo que a imunidade inata é fundamental para que se
desenvolva posteriormente uma imunidade específica contra determinado antígeno.
Por outro lado, a resposta secundária da imunidade específica utiliza inúmeros
componentes da imunidade inata para que essa resposta secundária específica
seja realmente mais rápida e eficaz.
Dentro da imunidade específica
pode haver dois tipos de imunidade que chamamos imunidade específica humoral e
imunidade celular. A imunidade humoral é mediada por anticorpos
produzido pelos linfócitos B, que têm como função neutralizar a infectividade dos agentes e sinalizá-los para a eliminação
através de vários mecanismos. Alguns anticorpos induzem a fagocitose por
células fagocíticas e outros desencadeiam a liberação
de mediadores da resposta inflamatória a partir de mastócitos.
A imunidade celular é mediada por linfócitos T que normalmente dirigem-se
contra células infectadas. Isso pode acontecer quando algum vírus, bactéria ou protozoário infecta e se prolifera dentro de fagócitos.
Para otimizar essa
interação entre os componentes do sistema imune, os linfócitos e as células
assessórias do sistema imune encontram-se localizados e concentrados em
órgãos denominados anatomicamente como órgãos linfóides primários e
secundários. Os primários são aqueles onde os linfócitos primeiramente se
desenvolvem, expressam os receptores de antígenos e atingem sua maturidade
funcional. São eles o timo (desenvolvimento de linfócitos T) e a medula óssea
(desenvolvimento de linfócitos B). Os órgãos imunes secundários são aqueles
para onde migram essas células B e T e onde ocorre a resposta destas aos
antígenos. São eles o baço, linfonodos, sistema imune
cutâneo e sistema imune de mucosas. O sistema imune de mucosas é muito importante
para conferir proteção contra patógenos que
tipicamente invadem o organismo através de suas superfícies.
No caso da utilização de vacina por via oral,
sabe-se que ao imunizar esta mucosa desencadeia-se uma proteção também em
outras mucosas(22). Outro ponto interessante neste sistema imune de mucosa são
as alterações morfológicas descritas por alguns pesquisadores. De acordo
com Pluske et
al. (1997) e Brown et al. (2006), em suínos, logo
após o desmame o intestino tem papel fundamental na defesa do organismo e por
isso aumenta sua taxa de renovação celular na membrana, alterando a relação vilo/crypta e aumentando a
atividade das enzimas digestivas na membrana em escova da mucosa
intestinal.
Desta maneira, fica claro que o sistema imune tem
papel importante também no desempenho animal podendo interferir no
funcionamento de outros órgãos e sendo interferido por diversos fatores,
principalmente nutricionais, como é o caso da troca alimentar e a presença de micotoxinas na dieta.
Micotoxinas
e sua interferência no sistema imune de suínos
Micotoxinas são
substâncias tóxicas produzidas por fungos que podem ocorrer em cereais
utilizados na ração animal e que causam inúmeras alterações no organismo tendo
principal influência no sistema imune. Acredita-se que 25% dos cereais
produzidos no mundo estão contaminados por micotoxinas.
Estudos indicam que as micotoxinas podem ser formadas
em várias partes da cadeia de produção alimentar como nos cereais ainda na
lavoura, durante o armazenamento, a fabricação e transporte de rações e durante
o armazenamento deste alimento nas granjas de produção animal(20).
Até hoje foram descritos mais de 300 diferentes
tipos de toxinas produzidos por distintas espécies fúngicas
e que podem afetar de diversas maneiras o sistema de defesa do organismo a
agentes estranhos com imunossupressão ou imunoestimulação.
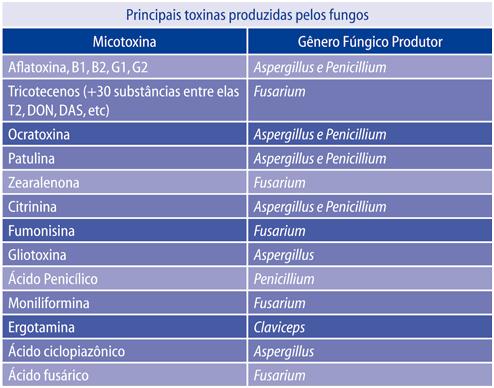
Na tabela 1 estão descritas algumas das principais toxinas produzidas pelos
fungos.
A maioria das micotoxinas
é descrita como inibidoras da síntese de proteí-nas
como DON, aflatoxina, ocratoxina
e muitos descrevem que a maior influência destas toxinas estaria relacionada ao
fato de diminuírem a cascata de síntese protéica tão necessária para os
mecanismos de defesa do organismo que envolvem desde
mitose celular até produção de proteínas como anticorpos e linfocinas,
provocando imunossupressão no animal exposto a estas toxinas.
Entretanto, Pestka et al. (2004) descreveram que deoxynivalenol (DON) e outros tricotecenos
podem tanto ter efeito imunoestimulatório como imunossupressivo dependendo da dose, freqüência de
exposição e status imunitário. De acordo com aqueles autores, baixas doses de tricotecenos interferem com a expressão de citocinas e genes inflamatórios induzindo imunoestimulação, enquanto altas doses promovem apoptose de
leucócitos com concomitante imunossupressão. Aquele estudo demonstra que
as micotoxinas podem também causar desregulação na sinalização celular com conseqüentes
alterações na expressão gênica de distintos tipos celulares.
Um estudo in vitro
realizado com células imunes humanas demonstrou que T-2 e fusarenon
X causam efeito imunossupresivo em linfócitos. Nivalenol e DON inibem a mitogenese
de células NK e zea-ralenona, alfa-zearalenol
e beta-zearalenol apresentam um efeito de inibição na
mitogenese de linfócitos somente em altas doses(3).
Um estudo realizado por Silvotti et al (1997) demonstrou que
quando porcas foram alimentadas com 800 ppb de aflatoxina durante gestação e lactação, foi detectado 500 ppt de aflatoxina M1 no leite e
os leitões nascidos destas porcas apresentaram-se alteração na imunidade
celular. No caso da aflatoxina, o mecanismo
responsável pela imunossupressão está relacionado com sua inibição na síntese
protéica.
Tanto a imunidade inata como a adquirida pode ser
afetada pelas micotoxinas. O efeito das micotoxinas pode ser ilustrado sobre o epitélio intestinal,
por exemplo, já que essa mucosa é considerada uma barreira física contra patógenos - possuindo ao mesmo tempo componentes da
imunidade inata quanto específica - como é o caso da presença de linfócitos e IgA. Na realidade, logo da
ingestão de um alimento contaminado, as células da mucosa intestinal podem
ficar expostas a grandes concentrações desta toxinas(19).
Como é descrito por Bouhet e Oswald (2005), a função
de barreira física realizada pelo epitélio intestinal é conseguido através da
resistência elétrica trans-epitelial (TEER) que existe na monocamada celular.
Algumas toxinas são capazes de diminuir essa TEER em células do intestino de
humanos como ocratoxina, patulina
e deoxivalenol. Bouhet et al., (2004) descreveram que fumonisina altera essa resistência trans-epitelial nas
células do intestino de suínos.
De acordo com aqueles autores, essa
pode ser uma
explicação dos processos de injúria,
descamação e ulceração observada em animais
expostos à ingestão de micotoxinas. Vários
investigadores procuraram explicar qual o mecanismo que as micotoxinas
poderiam afetar essa TEER na mucosa intestinal. McLaughlin
et al. (2004) explica que
isso pode acontecer devido a diminuição na quantidade de proteínas nas junções
celulares. De acordo com Leung et
al. (2003) a diminuição na biossíntese de esfingolipidios que é inibida pelas toxinas pode alterar a
regulação elétrica das células epiteliais.
Por outro lado, as células da mucosa intestinal que fazem essa proteção física inata são um tecido que está em
constante renovação para manter a integridade deste epitélio, o que ocorre a
partir da proliferação de células indiferenciadas provenientes da cripta e que
se diferenciam e se deslocam ao longo das vilosidades, sendo eliminados por
extrusão no ápice do vilo intestinal. Micotoxinas como T2 (Li et al, 1997) e ocratoxina(2) induzem necrose das células epiteliais da
cripta chegando à atrofia de vilosidades. Fumonisinas
são descritas como bloqueadora das fases G0/G1 do ciclo das células epiteliais
diminuindo sua proliferação(4). Baixas doses de DON interferem com a
diferenciação de enterócitos(10).
Ainda no que se refere à imunidade inata da
mucosa intestinal, sabe-se que a produção de muco a partir das células
caliciformes (também chamadas células Globbet) tem
importante função como lubrificação e barreira protetora deste epitélio.
Sabe-se que quando a mucosa intestinal é “desafiada” ocorre incremento no
número destas células no intestino com aumento na produção de muco. Isto é
demonstrado no caso de fatores antinutricionais dos
cereais (Oliveira et al.,
2000). Entretanto, somente um estudo na literatura demonstra que fumonisina induz hiperplasia de células epiteliais da
mucosa intestinal de frangos de corte(6). Mais estudos neste aspecto são
necessários para verificar a influência desta e outras micotoxinas
sobre a proliferação de células caliciformes e a produção de muco.
Na imunidade específica de mucosa destaca-se o papel das imunoglobulinas e células imunes como
linfócitos T. Em suínos, assim como em outros mamíferos, 80-90% das
imunoglobulinas presentes nos fluídos intestinais, lágrimas e glândulas
parótidas são IgA(08). De acordo com revisão de Bouhet et
al. (2005), toxina T-2 suprime as células B nas placas de Peyer
mas o efeito direto sobre a produção de IgA não foi
investigado. Por outro lado, Petska et al. (2004) afirmam que outros tricotecenos como DON e nivalenol
provocam uma dramática elevação na produção de IgA em
ratos, a ponto de o acúmulo destes anticorpos causar nefropatia
e hematuria.
Outro aspecto afetado pelas micotoxinas
é a produção de citocinas pelas células intestinais
que desempenham papel fundamental no recrutamento de células inflamatórias para
defesa desta mucosa. Oswald et
al. (2003) descreveu que leitões alimentados com baixos níveis de fumonisina diminuem a expressão de IL-8 no íleo, sugerindo
que este fato pode ter grande influência na maior susceptibilidade à E. coli observada nestes animais
quando comparado ao grupo controle. Aqueles autores sugerem que esse
menor recrutamento de células inflamatórias ocasionado pela diminuição na
expressão de IL-8 se associa a ação desta toxina na redução de proliferação
celular e integridade da mucosa do intestino aumentado a susceptibilidade dos
animais a colonização bacteriana.
A interferência de micotoxinas
na defesa das mucosas também foi descrita sobre a funcionalidade de macrófagos
alveolares em suínos. Liu et
al. (2002) demonstrou que tanto fumonisina B1 quanto aflatoxina apresentam citotoxicidade
dose e tempo dependente para macrófagos alveolares de suínos, porém a partir de
alterações distintas uma vez que são metabólitos totalmente diferentes. Fumonisina B1 inibe a esfigosina
e esfigosina N-acetiltransferase
que são fundamentais para a rota “de novo” de síntese de esfingolipídios. Este
é descrito como um evento inicial da citotoxicidade
da fumonisina em células. Alguns estudos demonstram
que a toxicidade da fumonisina para macrófagos de
aves (Qureshi and Hagler, 1992) é maior que a observada em suínos(12),
sugerindo uma diferença entre as espécies animais quanto à sensibilidade a esta
micotoxina.

Por outro lado, o efeito biológico de toxicidade
celular das aflatoxinas inclui citotoxicidade
aguda que requer a ativação do 2,3 epoxy metabólito
pela monooxigenase, ativando ligações covalentes
entre aflatoxina epoxidos
com macromoléculas. De acordo com Liu et
al (2002) macrófagos de suínos possuem essa monooxigenase para ativar aflatoxina
B1, gerando citotoxicidade nestas células de forma 10
vezes mais potente que a fumonisina B1. Aqueles
autores relataram ainda que somente a fumonisina induz apoptose nestes macrófagos, mas ambas as toxinas
diminuem a atividade fagocitica destas células. A fumonisina atuaria ainda reduzindo a produção de citocinas por essas células o que diminui a regulação da
resposta inflamatória nestes tecidos. Pang et al. (1987) observaram
diminuição na capacidade fagocítica de macrófagos
alveolares e na resposta blastogênica de linfócitos
pulmonares em suínos que inalaram T-2.
Esta influência dos metabólitos tóxicos de fungos
na imunidade das mucosas pode afetar muitíssimo o desempenho animal, já que a
indução desta imunidade é muito importante para conferir proteção contra
diversos patógenos que tipicamente invadem essas
superfícies. Em adição, existe aparentemente uma interrelação
entre as distintas mucosas do organismo, o que permite que a estimulação de uma
superfície mucosa potencialmente induza proteção específica em outras
mucosas(22). Como a administração de vacina via oral também é uma rota bastante
prática e econômica de imunização dos animais, quando ocorre a interferência de
micotoxinas nesta resposta imune, o resultado da
vacinação pode ser afetado.
Estudos em ratos também apresentaram que o efeito
da fumonisina sobre a resposta à imunização com SRBC
(eritrócitos de carneiro) depende do momento da administração da toxina, sendo
seu efeito evidente somente se a imunização ocorrer concomitantemente com a
vacinação(23). Aqueles autores também declararam que somente os machos, e não
as fêmeas, tiveram sua imunidade humoral afetada pela fumonisina.
O efeito de micotoxinas
afetando a produção da imunidade a vacinações já foi descrita em 1978 por Cysewski et
al. demonstrando que dietas contaminadas com aflatoxina
diminuíam a resposta de suínos a vacinação contra Erisipela. De acordo com
Oswald (2006), alimentação de suínos com 8 ppm fumonisina não modifica
a concentração sérica total de imunoglobulinas mas diminui
significativamente a resposta específica a um modelo de antígeno como a vacina
de micoplasma. Aquela mesma autora
afirma que em estudos in vitro a fumonisina
inibe a proliferação celular e altera a produção de citocinas,
referindo-se especificamente ao aumento na síntese de IFN-y, uma citocina de linfócitos T helper 1
(Th1) envolvidas na mediação da resposta imune celular e ao mesmo tempo,
diminui a IL-4, citocina Th2, envolvida na
resposta imune humoral.
Na verdade, todas essas alterações descritas na
literatura comprovam que as micotoxinas alteram a
resposta imune dos animais, podendo interferir com a resposta vacinal deixando
os animais susceptíveis a infecções inespecífica ou mesmo
para aquelas as quais esses animais tenham sido vacinados. Cabe ainda
ressaltar que a maioria dos estudos citados relaciona a adição de micotoxinas purificadas a dietas de boa qualidade. Na
prática, quando há presença de micotoxinas na dieta,
em algum momento ocorreu o desenvolvimento de fungos neste cereal e/ou ração.
Normalmente, esse desenvolvimento fúngico causa
perdas nutricionais, além de poder haver várias toxinas presentes, cujo
sinergismo ainda não é conhecido. Nestas condições é bem possível que o efeito
do uso de cereais e/ou alimentos contaminados com fungos e micotoxinas
na dietas dos animais possa promover interferências no
sistema imune dos animais que ainda não são conhecidas, mas que provavelmente
esteja relacionada a imunossupressão.
Infelizmente, está bastante distante a solução
para o controle total das micotoxinas em alimento
animal, mas o emprego de um programa adequado de controle de
fungos e micotoxinas pode ajudar a prevenir ou
minimizar esses efeitos deletérios das micotoxinas.