S.O.S. Suínos
Informativo Técnico nº 70
SUINOCULTURA
Como alimentar porcas que
desmamam 30 leitões por ano
AUTOR: Dalton de Oliveira Fontes
(Professor Associado da Escola de Veterinária da UFMG); Luisa Pinto de Oliveira
Souza; Gabriel Moreira Salum
1. INTRODUÇÃO
A produção de 30 desmamados/porca/ano que há poucos
anos parecia improvável hoje é uma realidade em algumas unidades produtoras de
suínos. O sucesso de desmamar tantos leitões por ano implica, quase
invariavelmente, em grandes e constantes desafios em todo sistema de produção,
seja relacionado ao manejo, sanidade ou nutrição. Plantéis que convivem com
essa nova realidade, se por um lado, tendem a ser mais rentáveis e
competitivos, por outro, exigem permanente controle no gerenciamento de todas as
etapas do sistema produtivo.
A rentabilidade de um sistema de produção está
diretamente ligada à eficiência reprodutiva, sendo que na suinocultura ela é
medida através do número de leitões desmamados por porca por ano. As fêmeas
modernas possuem como principais características um elevado número de leitões
nascidos, alta produção de leite, exigências nutricionais elevada, baixo
consumo voluntário, capacidade de deposição rápida de altas quantidades de
tecido magro, menores reservas de gordura. Com a seleção genética buscando
maior crescimento de tecido magro, o potencial das matrizes foi influenciado e
houve diminuição do consumo voluntário de alimento e do teor de gordura (Aherne
and Williams, 1992; Close and Cole, 2000). De acordo com Foxcroft (2005), as linhagens
maternas contemporâneas possuem maior potencial para crescimento de tecido
magro e isto, certamente, está associado com alterações no metabolismo em
geral, sendo necessário reavaliar as exigências nutricionais e as técnicas de
manejo nutricional, para otimizar o aproveitamento de nutrientes por parte
dessas fêmeas modernas. Tem sido relatado que uns dos aspectos que mais
restringe a produtividade das fêmeas suínas moderna é o manejo nutricional
inadequado prescindido a elas. Dessa forma, se torna cada vez mais importante a
compreensão de suas exigências nutricionais de forma a maximizar o desempenho
reprodutivo das fêmeas modernas.
Obviamente que matrizes hiperprolíferas apresentam
exigências nutricionais mais elevadas e por isso necessitam de um programa
nutricional diferenciado, seja durante sua preparação, gestação ou lactação.
Portanto, quando a nutrição for insuficiente para suprir as necessidades
nutricionais desses animais, a sua reprodução poderá ser seriamente
comprometida.
Este trabalho tem o objetivo de discutir as
estratégias nutricionais das fêmeas suínas modernas para se conseguir desmamar
30 leitões/porca/ano. Na primeira parte, será dada ênfase às estratégias
nutricionais para se aumentar a prolificidade das fêmeas, enquanto, na segunda,
serão abordadas considerações sobre as exigências desses animais.
PARTE I - ESTRATÉGIAS NUTRICIONAIS PARA PREPARAR FÊMEAS MODERNAS PARA
PRODUZIR 30 DESMAMADOS/PORCA/ANO
2. NUTRIÇÃO DAS LEITOAS DE REPOSIÇÃO
A seleção inicial para uma taxa elevada de crescimento
resulta em uma população não apenas mais jovem à puberdade, mas também mais
leve e, portanto, com peso mais baixo à maturidade sexual (Almeida, 1999).
Desse modo, na maioria das marrãs, a idade na qual estão aptas geneticamente
para o início do ciclo reprodutivo será atingida bem após terem alcançado uma
taxa de crescimento mínima ou o peso corporal necessário para o início da
puberdade (Foxcroft et al.,
1996). É bem possível que animais com genótipos extremamente magros e taxas de
crescimento muito elevadas e pouco apetite devido à seleção para menor
deposição de gordura não consigam atingir uma boa relação gordura:músculo para
desencadear a maturidade sexual, resultando no aumento da idade à puberdade
(Foxcroft et al., 1996). O
limiar de taxas de crescimento precisa assegurar que o crescimento per se não limite o início da maturidade
sexual da marrã (Foxcroft et al.,
2005).
A correta nutrição de marrãs durante o seu
crescimento tem impacto significativo no desenvolvimento reprodutivo das
fêmeas. Se for respeitada a utilização de níveis nutricionais e padrões de
consumo que permitam a ingestão de nutrientes em quantidade suficiente para
permitir taxas de ganho de peso comercialmente aceitáveis, bem como taxas de
deposição protéica compatíveis com as linhagens modernas, dificilmente
ocorrerão influências nutricionais relevantes sobre a idade à puberdade
(Almeida, 1999). Dessa forma, taxa de crescimento e composição corporal não
seriam limitantes à maturidade sexual (Foxcroft, 1993).
2.1. Nutrição das leitoas nas fases iniciais de crescimento
O manejo nutricional desses animais deve começar
nos estágios iniciais de sua vida. Na verdade, existem evidências que a
nutrição de leitoas no período uterino pode ter impacto na sua produtividade
subseqüente. Tem sido documentado, por exemplo, que a nutrição pode afetar o
desenvolvimento de fibras musculares dos fetos o que poderia ter impacto na
vida reprodutiva posterior. Desse modo, é importante estabelecer um adequado
programa de nutrição para evitar mudanças significativas na composição corporal
das matrizes em gestação.
Do nascimento até 3 semanas de idade, a leitegada
recebe em quantidade satisfatória os nutrientes necessários através do colostro
e do leite da porca. Pesquisas mais recentes indicam que o colostro, além de
imunoglobulinas, é rico em importantes fatores nutricionais de crescimento.
Leitões mais pesados à desmama têm melhor apetite durante a fase de creche e
crescem mais rapidamente do que leitões pequenos e mais leves. Por isso, ao
selecionar as marrãs de reposição, é preciso que os animais tenham nascido de
leitegadas mais pesadas, apresentem boas condições físicas, e, no mínimo, 6
pares de tetas funcionais. Além disso, há evidências sugerindo que leitoas
selecionadas de leitegadas pequenas (dez ou menos) atingem a puberdade mais
precocemente e por isso apresentam melhor desempenho reprodutivo subseqüente.
(Dritz, 2006).
Para a fase de creche, há limitada informação sobre
o efeito da nutrição sobre o desenvolvimento e longevidade das leitoas. Uma boa
estratégia de alimentação aliada a um ambiente controlado assegura a boa
qualidade das leitoas ao final do período de creche. Do mesmo modo, após a
saída de creche, para os grupos dos quais serão selecionadas as marrãs de
reposição, é necessário algum esforço quanto aos cuidados e ao manejo. Para
otimizar a alimentação de marrãs, aos produtores é recomendado algumas atitudes
simples como utilizar de comedouros e bebedouros compatíveis com cada fase
(crescimento ou terminação) e evitar alimentar as marrãs no chão devido ao
desperdício de ração e ao aproveitamento insuficiente da dieta.
O programa nutricional da fase inicial até a
puberdade tem como objetivo proporcionar às fêmeas uma adequada produção de
massa corporal (proteína) e deposição de gordura limitada e não o máximo ganho
de peso que é alvo dos programas de nutrição para animais em crescimento e
terminação destinados à produção de carne. Desse modo, é importante considerar
as marrãs como animais diferenciados dentro do plantel. Na tabela 1, são apresentadas
as especificações nutricionais para o desenvolvimento de marrãs segundo PIC
(2003).
Além disso, Dritz (2006) recomenda, para as dietas
de desenvolvimento de leitoas, que os níveis de cálcio e fósforo devam ser
0,10% mais altos daqueles recomendados para os animais destinados ao abate para
aumentar a densidade óssea e longevidade dos animais. Já os níveis de
suplementação de vitaminas e minerais devem ser semelhantes aos utilizados nas
dietas de suínos na fase crescimento.
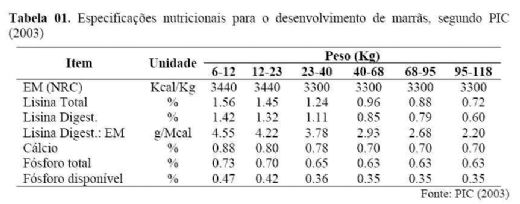
As variações observadas na literatura para as
especificações nutricionais para o desenvolvimento de marrãs ocorrem por
diversos fatores, sendo provavelmente a genética o mais importante. Assim, esse
deve ser um dos principais alvos a serem atingidos e será um ponto decisivo
para o sucesso reprodutivo do plantel. As tabelas e dados dos manuais da
genética devem, portanto, ser utilizadas como referência.
2.2. Nutrição das leitoas da seleção final à cobertura
Para
situações em que a seleção de leitoas de
reposição é feita com animais no final da
terminação o programa de alimentação
e manejo das leitoas, preferencialmente, deve ser baseado na idade e na
espessura de toucinho à época da seleção.
Após a seleção de marrãs, a taxa de
crescimento esperada para esses animais é de 650 g/dia
distribuídos em 450
g/dia de carne e ossos e
Por outro lado, leitoas selecionadas mais leves,
com maior espessura de toucinho e que permanecerão por um período prolongado de
reposição, antes da cobrição, devem receber uma dieta restrita ou com menor
densidade energética e que proporcionem um crescimento mais lento.
Entre as dificuldades em adequar o programa
nutricional das marrãs destacam-se: a falta de informações sobre as exigências
nutricionais de algumas genéticas; a utilização de diferentes genéticas dentro
do mesmo sistema de produção; a ausência de um manejo alimentar bem preparado e
executado até a primeira cobertura; e a falta de treinamento da mão-de-obra
nesta importante fase da produção. É importante salientar que algumas práticas
de manejo afetam o consumo de ração, como, por exemplo, a movimentação de
animais, a mistura de ninhadas diferentes e o estresse social, prejudicando a
execução.de um bom programa nutricional.
2.3. Nutrição das leitoas no período de
pré-cobrição (Flushing)
A nutrição específica de marrãs no período que antecede
sua primeira cobertura ou inseminação já é prática consolidada em boa parte da
indústria suinícola, uma vez que alguns autores já demonstraram que altos
níveis de energia nesta fase estão relacionados com a melhoria no desempenho
reprodutivo (Beltranena et al., 1991; Rhodes, 1991; Murgas, 1994; Ashworth,
1999; Ferguson et al., 2003;
Brustolini et al., 2004). Esta
prática é conhecida e mundialmente descrita pela expressão "flushing nutricional". Todavia, a
compreensão sobre os efeitos de diferentes fontes de energia dietética nesta
fase ainda carece de consolidação científica. Considerando o papel relevante
que a insulina pode desempenhar nas interações entre a nutrição e a reprodução
de marrãs, pode-se especular que dietas que efetivamente promovam o aumento nos
níveis de insulina plasmática venham a representar uma importante ferramenta
para a melhoria da eficiência reprodutiva dos rebanhos.
Tendo em vista que os efeitos nutricionais sobre a
taxa ovulatória parecem ser dependentes da ação da insulina (Cox et al., 1987; Britt et al., 1988; Matamoros et al., 1991), a manipulação da resposta
insulínica através da dieta pode representar um mecanismo de potencialização do
desempenho reprodutivo em suínos (van den Brand, 2001). Diversos outros autores
também sugeriram ou evidenciaram que a insulina pode intermediar os efeitos
interativos entre a nutrição e a reprodução de suínos.
2.4. Nova abordagem para o flushing com o objetivo de aumentar número de
nascidos
Face aos conhecimentos anteriormente expostos e às
hipóteses levantadas, Machado (2005) realizou um trabalho pela Escola de
Veterinária da Universidade Federal de Minas Gerais, com o objetivo de avaliar
o desempenho reprodutivo de marrãs cíclicas quando da utilização de rações
isoenergéticas baseadas em duas diferentes fontes de energia dietética. O
princípio básico para a definição dos tratamentos experimentais foi
fundamentado na hipótese que uma das fontes energéticas (carboidratos)
promoveria uma maior resposta insulínica aguda quando comparada à outra fonte
testada (lípides). Até a conclusão do referido trabalho, a hipótese só havia
sido experimentalmente testada em porcas lactantes (Kemp, 1995; van den Brand,
2001), as quais constituem um modelo experimental catabólico e, portanto, em
balanço energético negativo. As marrãs pré-cobertura, ao contrário das porcas,
representam um modelo experimental essencialmente anabólico. Essa diferença
marcante entre marrãs e porcas lactantes justifica a possibilidade da
ocorrência de respostas metabólicas distintas quando tais categorias são
submetidas às manipulações dietéticas propostas. Os trabalhos foram realizados
nas instalações da Granja Recanto, localizada no município de Patos de Minas.
Experimento 1 - Efeitos de diferentes fontes de energia sobre a eficiência
reprodutiva de marrãs cíclicas - I: efeitos sobre
taxa ovulatória, fertilidade, sobrevivência embrionária, morfometrias
embrionária e uterina, dinâmica ovariana e características do estro
Neste trabalho foram abordados os aspectos
referentes aos efeitos das diferentes fontes de energia sobre parâmetros de
desempenho reprodutivo, sobre a morfometria embrionária, uterina e placentária,
bem como às características do estro e da dinâmica folicular.
Após a primeira triagem e seleção, um grupo de 64
marrãs híbridas comerciais de uma mesma linhagem (cruzamento Landrace x
Large-White), oriundas de um lote de seleção contemporâneo, cujas idades
variavam entre 130 e 139 dias, com peso médio de
Durante a fase experimental propriamente dita,
todas as marrãs foram submetidas à avaliação quinzenal de peso, espessura de
toucinho e profundidade de lombo, além da determinação diária do consumo
individual.
No oitavo dia subseqüente ao dia do segundo estro,
de cada leitoa, iniciaram-se as dietas experimentais, que compunham os dois
tratamentos:
- Tratamento
1 (Grupo Flushing-Carboidratos):
marrãs alimentadas à vontade, entre o dia 8 do terceiro ciclo estral e o
cio subseqüente, com dieta cuja fonte adicional de energia é
predominantemente constituída por carboidratos (20% amido de milho).
- Tratamento
2 (Grupo Flushing-Lípides):
marrãs alimentadas à vontade, entre o dia 8 do terceiro ciclo estral e o
cio subseqüente, com dieta cuja fonte adicional de energia é
predominantemente lipídica (óleo refinado de soja, em quantidade exata
para substituir a energia fornecida pela fonte de amido incluída no
Tratamento 1, completando-se a diferença quantitativa com ingrediente
inerte).
As dietas eram isocalóricas, isoproteícas e
isolisínicas, sendo também constituídas pelas mesmas matérias-primas básicas
(milho, farelo de soja e farelo de trigo) e foram formuladas segundo Rostagno et al. (2000). O consumo ad libitum foi monitorado de forma a
avaliar se a ingestão diária de energia metabolizável seria tanto similar
quanto possível, para ambos os tratamentos.
Não houve efeito dos tratamentos sobre a taxa de
concepção (ou taxa de prenhez) após a inseminação artificial (tabela 3).
Considerando que, em espécies multíparas, o desafio metabólico mais
significativo está na quantidade e qualidade dos embriões concebidos, e não na
simples prenhez ou concepção em si, a situação fortemente anabólica das marrãs
deste experimento não justificaria diferenças nas taxas de concepção. A
diferença numérica encontrada entre os tratamentos, quanto à taxa de prenhez
(92,00% x 84,62%; não significativo, p>0,05), foi considerada como
casualidade experimental. Quanto aos diversos parâmetros indicativos do
desempenho reprodutivo, os resultados verificados ilustram uma significativa
relação entre o tratamento dietético aplicado e as diversas respostas
reprodutivas mensuradas. Em particular, a atividade ovariana parece ter sido
fortemente influenciada pelos tratamentos, sendo que o tratamento 1 (flushing com amido) demonstrou, de forma
geral, resultados significativamente superiores sobre o desempenho reprodutivo
das fêmeas, quando comparado com o tratamento 2 (flushing com óleo). A dieta flushing baseada em amido de milho aumentou a taxa
ovulatória (p=0,0052), o peso total dos ovários (p=0,069), o número total de
embriões viáveis (p=0,007), o comprimento médio dos embriões (p=0,069), o peso
médio dos embriões (p=0,002), a área média placentária (p=0,07) e o peso médio
das placentas (p=0,012). Neste trabalho a fonte de energia utilizada na ração flushing não influenciou a mortalidade
precoce dos embriões. Entretanto, não foi encontrado efeito da leitegada de
origem ("litter effect")
sobre a taxa de ovulação e sobre algumas características placentárias.
Tabla 03. Peso total do
útero gravídico (PTUG), número total de corpos lúteos (NTCL), peso total dos
dois ovários (PTO), número total de embriões (NTE), comprimento médio dos
embriões (CME), peso médio dos embriões (PME), área média da placenta por
embrião (AMP), peso médio da placenta por embrião (PMP), em marrãs cíclicas
submetidas aos tratamentos com amido de milho (T1) e com óleo vegetal (T2), com
seus respectivos coeficientes de variação (CV)
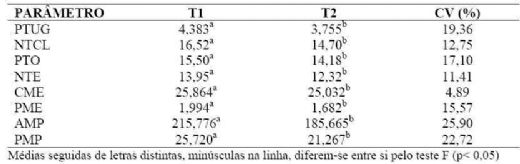
Experimento 2 - Efeitos de diferentes fontes de energia sobre a eficiência
reprodutiva de marrãs cíclicas - II: efeitos sobre a glicemia e as
concentrações séricas de insulina e progesterona
O presente trabalho enfoca os efeitos da onte de
energia dietética tanto sobre a curva glicêmica e a secreção insulínica, mas
também os possíveis efeitos sobre a produção de progesterona na fase
pós-ovulatória.
De um total de 54 marrãs selecionadas ao final da
fase pré-experimental e alocadas para os dois tratamentos, 21 unidades
experimentais foram aleatoriamente escolhidas para que fossem submetidas ao
procedimento de cateterização, de tal forma que estivessem igualmente
distribuídas entre os tratamentos dietéticos a serem aplicados no experimento.
O procedimento foi realizado sempre entre o quarto e o sexto dias do terceiro
ciclo estral (ciclo anterior ao da inseminação artificial), para cada uma das
marrãs utilizadas, de tal forma que a recuperação física e comportamental
necessária estivesse completa quando do início dos tratamentos nutricionais a
serem avaliados. Além das rotinas diárias já descritas, as marrãs
cateterizadas eram submetidas a um protocolo
contínuo de coletas de sangue, durante todo o período experimental. O protocolo
de coletas de sangue foi definido de acordo com as características e parâmetros
de secreção conhecidos para cada metabólito ou hormônio a ser avaliado. O
horário das coletas de sangue foi exatamente o mesmo horário definido para as
avaliações ultra-sonográficas da dinâmica ovariana. Para efeito de análise dos
dados, considerou-se que as coletas foram feitas com zero, seis, doze e dezoito
horas após a ovulação, lembrando que, neste caso, a hora-zero não significa o
momento exato da ovulação, mas sim o primeiro horário de coleta realizado após a
constatação da ovulação.
Os dados contidos na tabela 4 expressam a curva de
glicose no décimo quarto e vigésimo primeiro dia do ciclo estral, sete e
quatorze dias após o início dos tratamentos com as dietas experimentais.
Tabela 04. Concentração sangüínea de glicose, expressa em
mg/dL, no décimo quarto e vigésimo primeiro dia do ciclo de marrãs submetidas
aos tratamentos com amido de milho (T1) e com óleo vegetal (T2)

Verificou-se que não houve efeito dos tratamentos
sobre a glicose sangüínea, cabendo ressaltar que as marrãs já estavam há sete
dias sob influência metabólica da alimentação ad libitum de alta energia (flushing), utilizada neste experimento. Da mesma forma como
observado para o dia 14, não houve efeito de tratamento sobre os níveis de
glicose sangüínea, embora seja verificada uma diferença numérica relativamente
expressiva, mas não significativa (p=0,15) na coleta pós-prandial realizada aos
30 minutos após o fornecimento do alimento. Tanto no dia 14 quanto no dia 21 do
ciclo estral, as concentrações máximas de glicose sangüíneas foram detectadas
na coleta realizada aos 30 minutos após a ingestão do alimento matinal. Os
resultados obtidos no presente experimento sugerem ser a glicose um fraco
preditor das int
erações entre estado metabólico e reprodução, devido a sua estreita
regulação hormonal e relativa estabilidade, mesmo frente a distúrbios
metabólicos significativos. Entretanto, a presença de uma diferença numérica
expressiva (embora não estatisticamente significativa, p=0,15) permite-nos
especular sobre a possibilidade de resposta glicêmica potencialmente distinta
entre os tratamentos, principalmente em função da ascensão da curva observada
para o tratamento 1 (amido).
A hipótese principal testada neste experimento
envolveu também a determinação dos níveis de insulina sérica, avaliando o
padrão de secreção e os efeitos dos tratamentos sobre a resposta insulínica em
marrãs anabólicas. A tabela 5 contém os dados relativos à concentração sérica
de insulina no dia 14 do ciclo estral, sete dias após o início dos tratamentos.
Tabela 05. Concentração
sérica de insulina, expressa em μUI/mL, em função do tempo de coleta, no
décimo quarto e no vigésimo primeiro dia do ciclo de marrãs submetidas aos
tratamentos com amido de milho (T1) e com óleo vegetal (T2)

Ao contrário do que se observou para a glicose, já
se verifica, ao décimo quarto dia do ciclo, um efeito significativo (p<0,03)
dos tratamentos sobre a concentração sérica de insulina, mesmo com uma
quantidade reduzida de amostras para o tratamento 1 (n=6) e um coeficiente de
variação de 32,36%. As diferenças significativas ente os tratamentos
concentram-se nas duas primeiras coletas pós-prandiais, exatamente aquelas que
compõem o pico da resposta insulínica. Em função dos picos pós-prandiais serem
significativamente maiores (p<0,05) nas marrãs que recebem o flushing com amido, a média geral de
concentração de insulina nesses animais também é significativamente maior
(p=0,03) do que nas marrãs alimentadas com a dieta lipídica. Após 14 dias de
tratamento com as dietas isoenergéticas, o efeito das fontes de energia sobre
as concentrações médias de insulina e sobre seu pico pós-prandial permanece
significativos (p=0,067), se considerarmos a probabilidade do erro tipo 1 em um
limite de 7%.
A insulina parece estar fortemente envolvida nas
respostas observadas, seja por ação direta central e/ou gonadal, ou mesmo
indireta, através de fatores de crescimento locais intra-foliculares. As
diferenças significativas quanto à concentração sérica de insulina entre os
tratamentos concentraram-se nas duas primeiras coletas pós-prandiais,
exatamente aquelas que compõem o pico da resposta insulínica. Em função dos
picos pós-prandiais serem significativamente maiores (p<0,05) nas marrãs que
recebem o flushing com amido, a
média geral de concentração de insulina nesses animais também é
significativamente maior (p=0,03) do que nas marrãs alimentadas com a dieta
lipídica. Cabe salientar o efeito muito expressivo do tratamento sobre o peso
médio dos embriões aos 28,6 dias de gestação (1,994g vs
Um protocolo de coletas para determinação da
progesterona sérica foi implantado com o objetivo de avaliar a resposta
imediata de progesterona pós-ovulatória e associá-la com possíveis efeitos dos
tratamentos sobre a sobrevivência embrionária observada. Conforme pode ser
visualizado na tabela 6, não houve efeito dos tratamentos sobre a concentração
sérica de progesterona, avaliada até 18 horas após a ovulação estimada
Tabela 06. Concentração
sérica de progesterona, expressa em ng/mL, avaliada às 0h, 6h, 12h e 18h após a
ovulação, em marrãs cíclicas submetidas aos tratamentos com amido de milho (T1)
e com óleo vegetal (T2)

Nas condições em que foram realizados estes
experimentos, foi possível concluir que:
1) É possível alterar o padrão da curva de secreção
de insulina, bem como as médias de sua concentração sérica em marrãs, através
da substituição de uma fonte energética lipídica (óleo de soja) por uma fonte
baseada em carboidratos (amido de milho). Esta modificação do metabolismo
energético e da regulação hormonal pode ser conseguida mesmo em animais que
estejam em condições anabólicas e sem restrição alimentar, como o modelo
experimental testado neste trabalho;
2) A utilização de carboidratos como fonte
predominante de energia metabolizável na dieta flushing pode representar uma
eficiente ferramenta prática para a manipulação do metabolismo energético da
fêmea suína, induzindo efeitos anabólicos sobre o sistema reprodutivo e
otimizando a eficiência reprodutiva da espécie.
PARTE II - ESTRATÉGIAS NUTRICIONAIS PARA ALIMENTAR REPRODUTORAS SUÍNAS QUE
DESMAMAM 30 LEITÕES/ANO
3. NUTRIÇÃO DE PORCAS GESTANTES
Segundo Abreu et al. (2005), um programa nutricional para porcas gestantes
deve levar em consideração os seguintes aspectos: a) as diferentes fases e fenômenos
metabólicos que acontecem na gestação; b) as diferenças de padrão de
crescimento entre as porcas, segundo a ordem de parto; c) o estado metabólico
da matriz após a lactação anterior. Todos esses aspectos podem interferir nas
exigências nutricionais desses animais e devem ser consideradas para o
desenvolvimento de um programa nutricional. Matrizes hiperprolíferas apresentam
exigências nutricionais diferenciadas principalmente no terço final de gestação
onde ocorre o maior desenvolvimento das glândulas mamárias e dos fetos.
A nutrição durante a gestação além de influenciar o
desenrolar da gestação, o tamanho, o peso e a uniformidade da leitegada, afeta
também a produtividade no período da lactação, o intervalo desmama-cio e a
longevidade da porca. No primeiro terço da gestação, as necessidades
nutricionais são ligeiramente superiores às necessidades de mantença. Porém, o
fornecimento excessivo de energia nessa fase resulta em maior mortalidade
embrionária visto que o fluxo sanguíneo hepático aumenta e, com isso, a taxa de
metabolização da progesterona
também aumenta. A progesterona em menor
concentração no plasma sangüíneo influencia a baixa secreção de Proteína
Uterina Específica, reduzindo a taxa de sobrevivência embrionária (Sobestiansky
et al., 1998). Por outro lado,
Abreu et al. (2005), comentam que a condição corporal ou status energético da porca influencia a
respostas desses animais a altos níveis de consumo alimentar, sendo que a
mortalidade embrionária somente é aumentada em animais com boas condições
corporais.
No segundo terço da gestação o principal objetivo
de um programa nutricional é garantir o desenvolvimento corporal das fêmeas em
crescimento e a recuperação das condições corporais das matrizes, devido a
mobilização na lactação anterior. Nesse período o acompanhamento permanente da
condição corporal dos animais é de extrema importância.
Como comentado anteriormente o terço final de
gestação é o período em que há o maior desenvolvimento fetal e das glândulas
mamárias. Obviamente matrizes que crescem mais e que produzem mais leitões
apresentam um aumento nas exigências nutricionais de proteína, energia e
minerais.
3.1. Exigência de energia para porcas gestantes
Segundo Noblet et al. (1990), as exigências
nutricionais de uma porca gestante correspondem ao somatório das exigências
para mantença, crescimento uterino e ganho materno. Equações são usadas para
determinar a exigência de energia para mantença, ganho de peso materno e ganho
fetal. Resultados dessas equações, que foram convertidas em exigência diária,
estão representados na tabela 7.
Tabela 07. Exigência
de energia diária para matrizes com
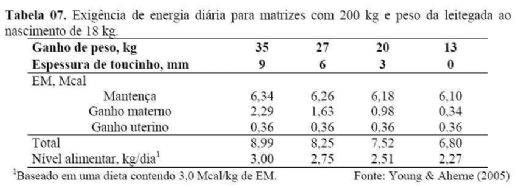
Pode-se observar que a exigência de energia para
mantença varia de
A exigência diária para porcas em gestação varia de
3.2. Exigência de proteína e lisina
para porcas gestantes
Segundo Kim e Wu (2005) fêmeas em gestação
necessitam de proteína para mantença, crescimento e crescimento dos conceptos
(fetos e membranas associadas). Desse modo, os aminoácidos da dieta devem estar
balanceados para garantir as necessidades dos fetos, das glândulas mamárias,
mantença e crescimento maternal. Segundo Close (2001) matrizes modernas em
crescimento exigem
As diferenças de exigências de aminoácidos e
energia entre pluríparas e fêmeas em crescimento deixam claro que os programas
de nutrição desses animais devem ser distintos. Por outro lado, Kim e Wu (2205)
propõem níveis e balanço de aminoácidos diferenciados para fêmeas de
3.2.1. Efeitos da suplementação da arginina durante gestação
A Arginina (Arg) desempenha múltiplos papéis no
metabolismo animal servindo de substrato para a síntese de proteína, como
intermediária no ciclo da uréia e como precursora na síntese de vários
compostos metabólicos importantes, incluindo o óxido nítrico (ON) e poliaminas
(Wu & Morris, 1998). O óxido nítrico é o maior vasodilatador das células
endoteliais (Wu & Meininger, 2000), e desempenha um papel importante na
regulação do fluxo sanguíneo placentário e, portanto, na transferência de
nutrientes e oxigênio da mãe para o feto (Bird et al., 2003). A Arg é também um
potente secretor de hormônios (Newsholme et al., 2005). Evidências crescentes
mostram que o ON e as poliaminas são chaves regulatórias da angiogênese e
embriogêneses, como também do crescimento placentário e fetal (Reynlds &
Redmer, 2001 e Wu et al., 2004a).
Perdas embrionárias e fatais devido a condições
intra-uterinas desfavoráveis durante a gestação representam o maior obstáculo
para a maximização da eficiência reprodutiva em animais de criação (Wu et al., 2006).
Como o maior fator que influencia no ambiente intra-uterino, a nutrição materna
desempenha um importante papel na regulação do crescimento, desenvolvimento e
sobrevivência fetal (Wu et al., 2004). Assim, fornecendo a porca gestante os
nutrientes adequados, incluindo quantidades adequadas de aminoácidos, é vital
para o crescimento fetal (McPheterson et al., 2004). A arg não é somente
exigida para a síntese de proteína e detoxificação da amônia, mas também é uma
precursora metabólica de muitas moléculas incluindo a prolina, ornitina,
poliaminas e ON (Wu & Morris, 1998; Kim et al., 2007). Além disso, a Arg é
a maior carreadora de nitrogênio para os fetos suínos e é um dos aminoácidos
mais abundantes nos tecidos fetais (Wu et al., 1999) e no fluido alantóide
durante o início da gestação (Wu et al., 1996), demonstrando a sua importância
na sobrevivência, crescimento e desenvolvimento fetal de leitões.
Mateo et al. (2007), realizaram um estudo para
verificar a hipótese de que aumentando o fornecimento de L-arginina aumentaria
o desempenho reprodutivo de marrãs prenhas entre o 30º e 114º dias de gestação.
Seus resultados indicaram que a suplementação com 1,0% de arginina-HCL entre os
dias 30 e 114 de gestação, aumentaram a concentração de arginina, ornitina e prolina
no plasma em 77, 53 e 30%, respectivamente. O tratamento com arginina não
afetou o peso corporal e a espessura de toucinho nas marrãs, mas aumentou o
número de suínos nascidos em 22% e o peso total da leitegada ao nascimento dos
leitões nascidos vivos em 24% (Tabela 1). A concentração plasmática de arg e
seus metabólitos (ornitina e prolina) foram alterados com o tratamento entre o
70º e 110º dias de gestação, o qual coincide com o período de rápido
crescimento fetal (McPheterson et al., 2004). A suplementação com arg reduziu o
número de natimortos em 65% (Tabela 1) provavelmente devido a uma melhora no
ambiente uterino para o crescimento e desenvolvimento fetal (Wu et al., 2006).
Tabela 08.Desempenho
reprodutivo de marrãs alimentadas com dietas com ou sem suplementação de 1%
L-arginina HCL

Semelhantemente, outros pesquisadores relataram que
suplementando 1% de arginina na dieta de fêmeas suínas entre os dias 14 e 28 de
gestação aumentou o número de leitões nascido em um leitão sem afetar a média
de peso ao nascimento (Remaekers et al., 2006). A suplementação durante a
gestação de porcas pode ter importantes aplicações para aumentar o desempenho
durante a gestação, como também o desenvolvimento pós-natal, a saúde e
qualidade de carne da progênie (Wu et al., 2006).
4. NUTRIÇÃO DE PORCAS LACTANTES
Durante a lactação das reprodutoras modernas as
exigências nutricionais apresentam-se bem altas já que as fêmeas contemporâneas
foram selecionadas para alta prolificidade e alta produção de leite. Aliado à
alta exigência nutricional, o consumo de alimentos normalmente é baixo. Assim
sendo, as fêmeas muitas vezes entram em balanço energético negativo e as
reservas corporais são prontamente mobilizadas, caracterizando o catabolismo
lactacional. A correta nutrição das matrizes durante a lactação também tem
impacto significativo no desenvolvimento reprodutivo subseqüente das fêmeas. Ao
respeitar a utilização de níveis nutricionais e padrões de consumo que
possibilitem a ingestão de nutrientes em quantidade satisfatória e a redução
dos efeitos do balanço energético negativo dificilmente ocorrerão influências
nutricionais relevantes sobre a eficiência reprodutiva desses animais.
Devido a mobilização de proteína corporal para
suprir as exigências da síntese de leite, o aumento da massa protéica corporal
da matriz ao parto parece proteger contra a falta de ingestão protéica na
lactação em porcas lactantes (Clowes et al.,
2003b). A perda de mais de 12% de massa protéica pela porca ao parto parece
reduzir significativamente a fertilidade destas fêmeas após a desmama.
É importante que os níveis nutricionais e de
consumo sejam respeitados dentro dos padrões de cada genética, pois mesmo
restrições modestas em estágios críticos do desenvolvimento folicular podem ter
implicações duradouras para a função reprodutiva das fêmeas no plantel. Além
disso, considerando que primíparas modernas foram selecionadas para alta
prolificidade e alta produção de leite e ao mesmo tempo apresentam capacidade
limitada de consumo durante a lactação, as exigências nutricionais desses
animais apresentam-se bem altas e por isso devem ser estabelecidos programas
nutricionais distintos das matrizes.
Na tabela 09, são apresentadas especificações
nutricionais na fase de lactação para primíparas e matrizes.
Recomendações nutricionais de marrãs e porcas durante a lactação
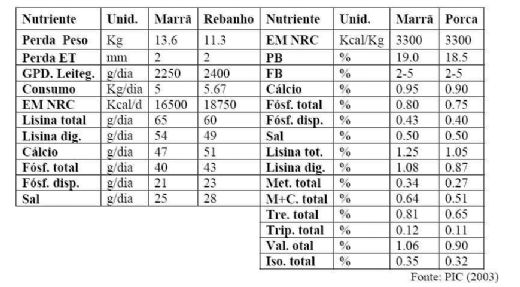
4.1. Exigência de energia total durante a lactação
A exigência de energia metabolizável total durante a lactação (EMlact)
corresponde ao somatório da exigência energética de mantença e a exigência para
a produção de leite. Segundo Noblet et al. (1998), a exigência total de energia
de uma porca durante a lactação pode ser representada pelas seguintes equações:
EMlact (MJ/dia) = 0,460 x peso metabólico da matriz (PC0,75) + Eleite/0,72,
a Eleitedurante toda a lactação pode ser estimada pela equação 3 da tabela 3.
EMlact (MJ/dia) = 0,460 x peso metabólico da matriz (PC0,75) + (28,6 x
ganho de peso diário da leitegada (kg) - 0,52 x tamanho da leitegada)/0,72
Ilustrações da aplicação da equação sugerida por Noblet et al. (1998) são
visualizadas na tabela 10 para fêmeas que desmamam 10, 11 e 12 leitões (fêmeas
melhoradas de alta prolificidade e alta produção de leite).Exigência diária de
energia metabolizável de porcas em lactação com
Tabela 10. Exigência diária de energia metabolizável de porcas em lactação
com
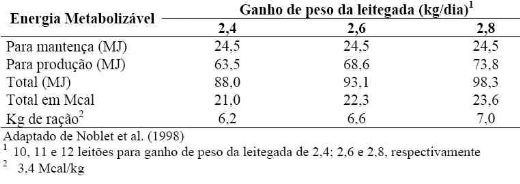
Os dados da tabela indicam que as exigências energéticas totais de porcas
em lactação são muito superiores do que as de porca em gestação e que podem, na
maioria das condições comerciais de criação, não serem atendidas pelo consumo
alimentar da porca. Além disso, observa-se que as necessidades energéticas para
matrizes que desmamam 12 leitões são aproximadamente 12% superiores em relação
a matrizes que desmamam 10 leitões, ou seja, para se produzir dois leitões a
mais a matriz precisa consumir cerca de
4.2. Exigência de proteína e lisina para porcas lactantes
Na tabela 13, são apresentadas estimativas de exigência de lisina para
porcas em lactação com
Pode-se observar que matrizes que desmamam uma leitegada de 12 leitões
(ganho de peso médio da leitegada de 2,8kg) exigem aproximadamente
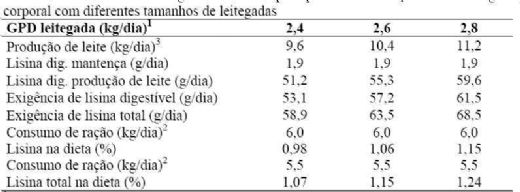
1 - 10, 11 e 12 leitões para 2,4, 2,6 e
2,8 kg/dia, respectivamente.
2 - Consumo estimado de ração por dia
3 - Segundo Close (2001):
Exigência de lisina para
mantença:
0,036 g x peso corporal(kg) 0,75
Exigência de lisina para produção de
leite:
Produção de leite (g/dia) = 4 x ganho
de peso da leitegada (g/dia)
Produção de proteína no leite (g/dia) = produção de leite x 0,056
Produção de lisina total (g/dia) = produção de proteína no leite (g/dia) x
0,076
Eficiência de utilização de lisina para produção de leite = 80%
Digestibilidade da lisina = 90%
Quando as porcas não recebem quantidades adequadas
de aminoácidos dietéticos, tecidos protéicos maternos são mobilizados
(especialmente proteínas do músculo esquelético). Mobilização excessiva de
tecidos protéicos maternos, muitas vezes, resulta em prejuízos reprodutivos nos
partos subseqüentes (Jones & Stahly, 1995). Portanto, estabelecer a
exigência nutricional de porcas em lactação não está apenas restrito à
maximização da produção de leite, mas estende também para a manutenção da
condição corporal da porca para os partos subseqüentes (NRC, 1998; Kim &
Easter, 2003). A mobilização protéica ocorre em vários tecidos da porca com
diferentes taxas (Escobar, 1998; Kim & Easter, 2001). O músculo é o maior
doador de aminoácidos durante a privação alimentar ou inadequado fornecimento
de proteína dietética, considerando que o trato reprodutivo contribui com a
maior porção do seu próprio aminoácido (Kim & Easter, 2001).
4.3. Efeitos da suplementação da Arginina durante a lactação
A Arginina (Arg) é um aminoácido essencial para os
leitões (Southern and Baker, 1993; Roth et al., 1995; Urschel et al., 2006),
devido a grande utilização da Arg pelas múltiplas vias metabólicas (Wu &
Morris, 1998; Li et al., 2007). Porém, o consumo de Arg através do leite da
porca é baixo em relação à exigência de proteína dos leitões (Davis et al.,
1994; Wu & Knabe 1994; Kim et al., 2007). Estimativas baseadas na
suplementação de Arg pelo leite da porca e na exigência de Arg dos leitões
revelaram que o leite da porca fornece menos que 40% da exigência diária de
leitões lactentes com 7 dias de idade (Wu et al., 2004).
Kim et al. (2004), relataram que a suplementação
com 0,2 e 0,4% de L-arginina para leitões lactantes de
O aumento da ingestão de Arg pelo leite pode ser
uma maneira efetiva de aumentar o crescimento dos leitões. Além de o consumo alimentar
da porca e da intensidade de mamada dos leitões, a produção de leite também é
influenciada pela angiogênese do tecido mamário e pelo fluxo sanguíneo para as
glândulas mamárias, o qual aumenta o fornecimento de nutrientes para as
glândulas mamárias para a produção de leite (Trottier et al., 1997). O fluxo
sanguíneo mamário e a angiogênese são regulados pela Arg derivada do óxido
nítrico (Meininger & Wu, 2002; Lacasse & Prosser, 2003). Além disso, a
produção de leite é altamente correlacionada com o crescimento da glândula
mamária (Kim et al.,2000) e a Arg é necessária para otimizar o crescimento da
glândula mamária (Pau & Milner, 1982). Em altas dosagens, a Arg estimula a
secreção de prolactina e hormônio do crescimento, os quais são necessários para
o desenvolvimento mamário (Knopf et al., 1968; Davis, 1972).
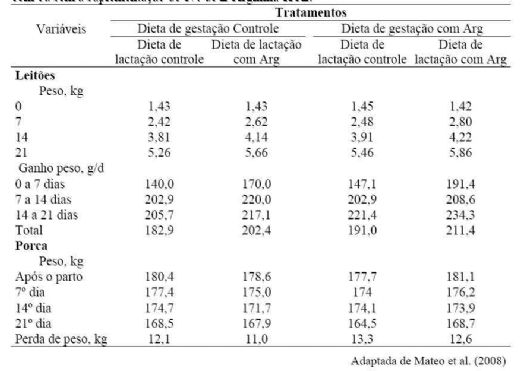
Mateo et al. (2008), sugeriram a hipótese de que a
suplementação de Arg em dietas de fêmeas de primeiro parto durante a gestação e
lactação pode estimular o ganho de peso dos leitões lactantes, possivelmente
pelo aumento da utilização de nutrientes e, conseqüentemente, aumentando a
produção de leite e modificando a composição nutricional do leite. Nesse
estudo, Mateo et al. (2008) demonstraram que a suplementação de Arg na dieta
das porcas durante toda a lactação aumentou a concentração total de aminoácidos
no leite e melhorou o desempenho de crescimento dos leitões. Essas descobertas
forneceram uma nova estratégia de manejo nutricional de porcas recém paridas.
Sabe-se que o ganho de peso da leitegada é
correlacionado com a produção de leite ou concentração de nutrientes do leite
(Kim et al., 1993). O aumento do ganho de peso do leitão o da leitegada em
dietas de porcas suplementadas com Arg pode ser um indicativo de aumento da
produção de leite ou aumento da concentração de nutrientes no leite. Resultados
do estudo de Mateo et al. (2008) indicam que o consumo voluntário e mudanças no
peso vivo das porcas não foram afetados pela dieta suplementada com Arg (tabela
2), sugerindo que aumentos na concentração total de aminoácidos (AA) no leite
não foram devidos a alterações no consumo alimentar protéico ou mobilização de
proteínas corporal. Baseado na redução dos níveis de uréia no plasma, a
suplementação com Arg parece aumentar a eficiência de utilização protéica para
a síntese de proteínas do leite. Pelo aumento da síntese de ON (o maior
vasodilatador) das células endoteliais dos vasos sanguíneos (Wu &
Meininger, 2000), a suplementação dietética de Arg pode aumentar o fluxo
sanguíneo e o fornecimento de nutrientes para as glândulas mamárias para a
produção de proteínas do leite, resultando em aumento do ganho de peso dos
leitões lactentes. O aumento da concentração total de AA no leite foi associado
com o aumento do ganho de peso dos leitões durante a primeira semana de
lactação, a qual afeta todo o desenvolvimento dos leitões durante toda a
lactação. Em média, leitões de porcas suplementadas com Arg ganham
Mateo et al. (2008) concluíram que a suplementação
dietética de Arg em porcas lactantes aumenta o desempenho dos leitões
lactentes. O aumento de ganho de peso da leitegada foi associado com o aumento
na concentração total de AA no leite no 7º dia de lactação. Os pesquisadores
sugerem que o tratamento com Arg pode aumentar o fluxo sanguíneo mamário e a
mobilização de AA durante a lactação. Porém, mais estudos ainda são necessários
para testar essa nova hipótese.
4.4. Exigência de outros aminoácidos para porcas lactantes
Os estudos para estimar a exigência de valina para matrizes lactantes
apresentam resultados contraditórios. Segundo Kim (2005), para porcas com baixo
consumo voluntário de alimento e uma mobilização tecidual substancial durante a
lactação (i.e., porcas primíparas e de segundo parto), a treonina é um aminoácido crítico,
enquanto a valina se torna crescentemente importante em porcas que têm um alto
consumo alimentar e uma limitada mobilização tecidual (i.e., porcas multíparas)
durante a lactação (Kim et al., 2001). O NRC (1998) propôs uma relação mínima
de valina:lisina (V:L) de 0,85:1 para matrizes de alta produção de leite. Esta
relação foi maior que a proposta previamente pelo ARC (1981) e por Pettigrew
(1993), mas é menor que a proposta pela edição anterior do NRC (1988). Richert
et al. (1996, 1997a,b) estimaram que a exigência de valina para porcas
amamentando ≥ 10 leitões foi maior que a estimada por Pettigrew (1993) e
pelo NRC (1988). Já Carter et al. (2000) e Gaines et al. (2006) relataram que a
exigência de valina para porcas prolíficas em lactação não excede os valores
estimados pelo NRC (1998).
A glutamina
aparece em grandes quantidades no leite de matrizes suínas,
culminando com uma redução pronunciada de glutamina na musculatura esquelética
de matrizes lactantes. Wu (2005) concluiu que uma suplementação durante a
lactação poderia prevenir a perda de massa muscular de matrizes suínas
gestantes. Complementando esta afirmação, Lobley et al. (2001) acrescentaram que a glutamina pode limitar a
produção de proteína no leite, particularmente durante o período inicial da
lactação, quando as concentrações de glutamato e glutamina diminuíram em torno
de 25%. Neste mesmo período, também ocorreu uma diminuição em torno de 25% de
glutamina no músculo das matrizes suínas.
Até o presente momento, o único estudo conhecido na
espécie suína, durante as fases de gestação e lactação, foi conduzido por Kitt et al. (2004), no qual estudaram o efeito
da suplementação com 2,5% de glutamina (Gln), na dieta da matriz suína, em
substituição ao farelo de soja. Como resultados encontraram 17% e 36% a mais
nas concentrações plasmáticas de glutamina no 7º e no 21º dia de lactação,
respectivamente, quando comparadas com as matrizes suínas mantidas com dieta
controle. Os autores acrescentaram ainda que as matrizes suínas suplementadas
com 2,5% de Gln no 7º e 21º dias de lactação obtiveram 46% e 265% a mais nas
concentrações de glutamina no leite, respectivamente, quando comparadas com
àquelas do grupo controle.
Manso (2006) avaliou as concentrações sanguíneas,
musculares e lácteas de glutamina em matrizes suínas primíparas submetidas a
uma suplementação com glutamina. Foram utilizadas 45 matrizes suínas primíparas
distribuídas em 3 tratamentos: T1 (controle), T2 (2,5% L-glutamina) e T3 (2,5%
AminoGut®). As matrizes iniciaram a suplementação 30 dias antes do parto
continuando até o desmame. Não houve diferença estatística para a concentração
de glutamina no músculo e no sangue dos grupos experimentais para os períodos
estudados (P>0,05). Com relação à concentração de glutamina no leite, no 7o
dia de lactação as leitoas do T2 apresentaram uma maior concentração de
glutamina enquanto que aos 21 dias de lactação as matrizes suínas primíparas do
grupo T3 apresentaram uma maior concentração de glutamina no leite (P<0,01).
Outros estudos deverão ser feitos para se avaliar diferentes níveis de
suplementação de glutamina associados a diferentes tamanhos de leitegadas para
avaliar o subseqüente desempenho reprodutivo das matrizes e o desempenho dos
leitões após o desmame.
5. EXIGÊNCIA DE MINERAIS PARA PORCAS COM O OBJETIVO DE DESMAMAR 30
LEITÕES/PORCA/ANO
Com a introdução de novas linhagens maternas
capazes de produzir leitegadas de tamanhos maiores, maior peso ao nascimento e
porcas com maior produção de leite,
resultando em mais leitões desmamados e maiores
pesos ao desmame, a demanda nutricional desses animais tornou-se excessivamente
alta. As recomendações minerais do NRC (1998) para suínos não mudaram
substancialmente nos 25 anos passados (
Para ajustar antecipadamente a maior necessidade
biológica para minerais pela alta produção das linhagens maternas, a indústria
de ração e os especialistas das universidades recomendam rotineiramente níveis
dietéticos mais elevados, reforçando os níveis dos macro e micro minerais, bem
como de outros nutrientes, nas dietas de gestação e lactação. Embora esta
prática seja lógica e possa ser exatamente o que essas porcas necessitam para
altas produtividades, geralmente não é baseada em pesquisas, mas em observações
do campo, ou seja, de forma empírica.
Cálcio e fósforo são os dois principais minerais
geralmente associados com a integridade e estrutura dos pés. As pesquisas de
Nimmo et al. (1981)
demonstraram que uma alta porcentagem de leitoas eram incapazes de chegar ao
primeiro parto quando alimentadas com dietas contendo os níveis de Ca e P
preconizados pelo NRC (1978) durantes os períodos de desenvolvimento e
reprodução. Outro trabalho demonstrou que porcas de alta produtividade têm
maior ocorrência de desmineralização óssea durante sua vida reprodutiva (Maxson
e Mahan, 1986). Estes resultados combinados sugerem que durante períodos de
crescimento e períodos reprodutivos há uma alta demanda por Ca e P. Estes dois
minerais podem, assim, ser removidos do tecido ósseo quando as demandas para
produção de leite são grandes.
Mahan (2006) publicou os dados sobre a avaliação da
deposição fetal de minerais, dos 45 dias após a cobertura ao terço final de
gestação. Os dados desse trabalho serão descritos a seguir.
O autor observou que o índice total de minerais do
corpo de leitegadas em desenvolvimento aproximadamente dobrou a cada
Ao parto, a concentração de Ca no colostro da porca
é baixa, mas aumenta com o progresso da lactação (Figura 2 - A). O baixo nível
de Ca ao parto pode ser explicado pela sua diminuição na porca gestante, pois
ela transfere grande quantidade de Ca a seus fetos
Em conseqüência dos resultados acima a deposição total de Ca e P nos
leitões desmamados (11 ou 21 dias) foi maior nas leitegadas maiores (Figura 1 -
III e IV). Quando expressado em uma base individual não houve nenhuma diferença
no teor de Ca e P nestes leitões, sugerindo que a progênie tem aproximadamente
os mesmos teor minerais.
Os
dados apresentados nas figuras 1- I e 2- A implicam que a maior
resposta
ao aumento da retenção mineral no desenvolvimento da
leitegada foi atribuída ao
Ca e P. Os dados demonstraram que todos os minerais essenciais seguiram
o mesmo
padrão geral da retenção como o Ca e o P, mas eles
demonstraram também algumas
diferenças. Esta diferença é atribuída
às diferentes funções biológicas de cada
elemento como quando este é necessário em um
estágio específico de
desenvolvimento do feto ou no terço final da
gestação, onde o feto retém
minerais para sua vida pós-natal subseqüente.
Dois dos elementos residuais críticos (Fe e Zn) serão apresentados na Figura
1 - V. Embora a quantidade de Zn não dobrasse durante as últimas 2 semanas do
desenvolvimento fetal, como fez Ca e P, a quantidade de Fe aumentou
extremamente durante este último período de gestação. Entretanto, embora seja
mostrado um grande aumento no teor total do Fe durante o terço final de
gestação, este ainda permanece abaixo daquele necessário para o recém nascido.
Conseqüentemente, uma fonte exógena é necessária para impedir a anemia no
neonato, uma vez que a quantidade de Fe secretada no tecido mamário é
considerada inadequada às demandas elevadas de Fe do suíno em rápido
crescimento. O colostro da porca e a composição do Fe no leite são apresentados
na Figura 2 - C.
O conteúdo de Fe no colostro e mais tarde no leite declina com o progresso
da lactação, e este seu teor durante lactação parece ser influenciado pelo
número de leitões lactentes. Os leitões de leitegadas maiores recebem um leite
com teor de Fe mais baixos do que aqueles animais nutridos pelas porcas em
leitegadas menores. O teor total de Fe em leitegada de 8 ou 11 leitões, quando
desmamados aos 11 ou 21 dias da idade, mostrou que aquelas leitegadas maiores
tiveram maiores conteúdos de Fe total (Figura 1 - IV). .
O teor do Zn da leitegada durante o desenvolvimento da gestação apresentado
na figura 1-V demonstra que o conteúdo do Zn na leitegada aumentou extremamente
durante o período da gestação com o maior aumento ocorrendo durante as últimas
2 semanas de gestação. O colostro teve concentração de Zn mais elevada quando
comparado ao leite (Figura 2- D), com a concentração do Zn declinando no leite
mais tardio e aparentemente há pouco efeito do tamanho da leitegada na
concentração de Zn no leite durante a lactação. O conteúdo total do Zn na
leitegada aumentou á medida que o leitão alcançou a idade de desmame, sendo que
as leitegadas maiores tiveram teores totais Zn mais elevados (Figura 1 - VII).
5.1. Fontes minerais orgânicas x inorgânicas
Os minerais executam diversos papéis no corpo e são essenciais para
diversas funções reprodutivas. Não somente estão envolvidos no controle da
enzima de vários processos metabólicos e hormonais, mas são igualmente
importantes nos processos de crescimento, saúde e controle imunológico. Quando
providos em excesso são retidos no fígado, mas tem sido mostrado que eles podem
ser pró-oxidantes e assim podem ser prejudiciais às funções corporais.
Conseqüentemente, a forma em que o elemento é fornecido ao animal pode
tornar-se mais importante no futuro com o aumento das necessidades dietéticas.
Por exemplo, o fornecimento de Se orgânico ou inorgânico, desde que respeitando
a exigência do suíno, realçará o controle anti-oxidantes e do sistema imune.
Entretanto, quando uma ou outra forma é fornecida em algum excesso, observa-se
que muito do Se orgânico é retido no tecido visto que foi mostrado que o Se
inorgânico adicional pode causar lesões oxidativas no tecido, sendo assim,
prejudicial ao desempenho animal. Com o aumento das necessidades minerais
das
porcas, há simultaneamente um interesse em aumentar níveis minerais nas
dietas durante a gestação e a lactação. Um recente estudo avaliou os níveis
minerais das porcas quando alimentadas com minerais inorgânicos (forma de
sulfato ou de óxido) ou orgânicos. O experimento incluiu níveis do NRC (1998) e
um alto nível tipicamente fornecido pela indústria. O experimento envolveu um
total de 375 leitegadas e os resultados são apresentados na Figura 3 (Peters,
2006).
Embora não significativo, o número total de leitões nascidos vivos foi,
aproximadamente, de um leitão adicional por leitegada quando porcas foram
alimentadas com minerais orgânicos. Como evidente na figura 3 não houve nenhuma
diferença no número de leitões nascidos vivos quando os níveis do NRC (1998)
foram fornecidos. Embora esta experiência precise ser confirmada em outros
animais, os resultados sugerem que os minerais orgânicos possam ser superiores
aos minerais inorgânicos, e que o reforço extra dos minerais na forma orgânica
pode ser benéfico ao desempenho reprodutivo da porca.
5.2. Status mineral
da porca
As porcas durante sua vida reprodutiva têm maior necessidade de minerais,
tendo uma perda maior de minerais do corpo durante a gestação e lactação (Mahan
e Newton, 1995). Aumentos de produtividade da porca são acompanhados pelo
aumento da demanda nutricional e, assim, reservatórios de minerais do seu corpo
poderiam diminuir. Quando os teores minerais das porcas completando 3 partos
foram comparados a leitoas que não reproduziram de mesma idade, os resultados
mostraram que a maioria dos minerais era mais baixa nas porcas que tinham se
reproduzido (Figura 4).
O Ca e P tiveram a maior perda, aproximadamente, com teores totais de
Embora as exigências minerais sejam mal definidas para animais de
reprodução (Hostetler e et al., 2003) é necessário continuar pesquisando para
determinar estas necessidades. Porcas de alta produtividade possuem maiores
necessidades de minerais do que porcas de baixa produtividade. Os resultados
apresentados implicam que há talvez "uma relação ideal" e uma
"janela crítica da necessidade" para os minerais para a reprodução.
Esta "relação ideal dos minerais" e suas necessidades biológicas em
períodos de tempo específicos durante a gestação podem igualmente ser
diferenciadas pelo estágio da reprodução. Animais de alta produtividade têm
exigência nutricional mais elevada para estes minerais, sendo assim, esgotam as
suas reservas corporais.
Embora
nós sejamos acostumados a aumentar os minerais dietéticos
em
proporção às necessidades estimadas, esta
prática pode ser um erro por causa
das "janelas de diferença da necessidade" e dos potenciais
efeitos
prejudiciais de níveis adicionais. As necessidades minerais
talvez sejam
reguladas geneticamente e pelo tamanho da leitegada, visto que as
secreções
mamárias da lactação não podem somente
refletir o padrão genético de secreção
do leite, mas podem igualmente refletir um caminho onde o excesso de
minerais
pode ser excretado pelo corpo.
As exigências dos minerais podem ser influenciadas pela forma do mineral
fornecido, sendo igualmente possível que minerais dietéticos mais elevados
podem ser desejáveis em alguns estágios da reprodução e prejudiciais em outros
estágios. O papel dos minerais orgânicos nesta área é até agora desconhecido,
mas resultados indicam que podem ter uma influência positiva no desempenho
reprodutivo da porca, visto que minerais inorgânicos podem ser prejudiciais
quando fornecidos a níveis "normais" mais elevados, particularmente
quando fornecidos continuamente. A determinação das necessidades minerais da
porca de reprodução está certamente incipiente quando comparada a de outros
nutrientes. Uma pesquisa mais extensiva precisa ser conduzida, visto que a
exigência dos animais parece ser aumentada com a capacidade genética dos
animais.
FIGURA 1Teor de minerais na leitegada (g): I) Teor
mineral total; II) Teor total de Cálcio e Fósforo; III) Teor total de Cálcio;
IV) Teor total de Fósforo; V) Teor total de ferro e zinco; VI) Teor total de
ferro; VII) Teor total de zinco .
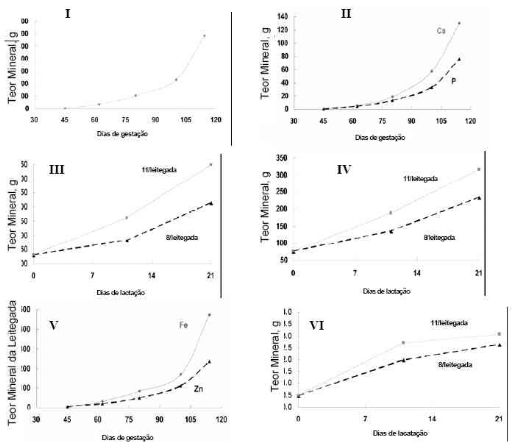
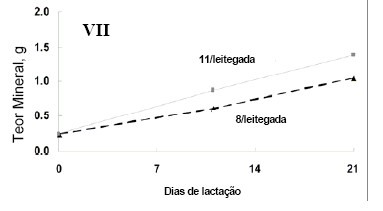
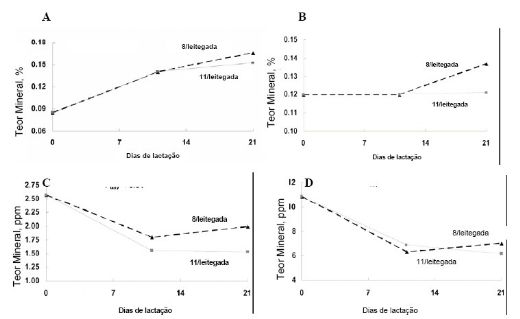
Figura 2. Teor de minerais no colostro e no leite ao nascimento, 11 e 21
dias pós-parto de porcas com dois tamanhos de leitegadas; A) Teor de Cálcio; B)
Teor de Fósforo; C) Teor de ferro; D) Teor de zinco.
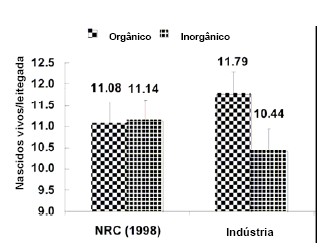
Figura 3. Efeito de fontes orgânicas e inorgânicas de minerais a dois
níveis dietéticos sobre 6 partos.
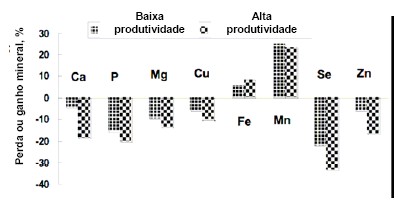
Figura 4.Perdas ou ganhos minerais após o terceiro parto (comparado ao
controle não prenhe)
6. CONSIDERAÇÕES FINAIS
Fêmeas suínas que desmamam em média de 12 leitões e que apresentam 2,5
partos/ano irão desmamar 30/animais/ano. Obviamente, essas reprodutoras
modernas além de alta fertilidade terão o desafio de criar bem os leitões, ou
seja, boa habilidade materna. Para alcançar e manter esse novo desafio um programa
nutricional de precisão se faz necessário, em todas as fases da vida. Nesse
sentido, a compreensão das exigências dos animais nas diferentes fases de
crescimento, na reposição, no flushing, na gestação (principalmente no terço
final) e na lactação se faz necessário.
Nos últimos anos muitas informações têm dado sustentabilidade aos
nutricionistas para o desenvolvimento dos programas nutricionais de matrizes
hiperprolíferas, porém novos desafios estão sendo diariamente lançados, e
pesquisas são necessárias para adequação permanente dessas estratégias
nutricionais.
7. REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, M.L.T.; DONZELE, J.L.; OLIVEIRA, R.F.M. Exigências nutricionais de
matrizes suínas gestantes e lactantes. In: IV Seminário Internacional de Av4es
e Suínos - Avesui, p.33-59, 2005.
ADASHI, E.Y. et al. Insulin
enhancement of luteinizing hormone and
follicle-stimulating hormone release by cultured pituitary cells. Endocrinology, v.108, p.1441-1449, 1981.
AHERNE, F.X., WILLIAMS, I.H. Nutrition for optimizing breeding herd
performance. Veterinary Clinical North American Food Animal Practice
8:589-608, 1992.
ALMEIDA, F.R.C.L. Interações entre nutrição e reprodução
ASHWORTH, C.J. et al. Effects
of pre- and post-mating nutritional status on hepatic function, progesterone
concentration, uterine protein secretion and embryo survival in Meishan pigs. Reprod. Fertil. Dev.,
v.11, n.1, p.67-73, 1999.
ARC. Agricultural
Technical Review. In The Nutrient Requirements of Pigs: Rev. ed. Commonw. Agric. Bureau,
BELTRANENA, E. et al. Effects of pre- and postpubertal
feeding on production traits at first and second estrus in gilts. J.
Anim. Sci., v. 69, p.886-893, 1991.
BORTOLOZZO, P.; WENTZ, I. A fêmea suína de reposição.
BRONSON, F.H. Mammalian reproductive biology.
BRITT, J.H.; ARMSTRONG, J.D.; COX, N.M. Metabolic
interfaces between nutrition and reproduction in pigs. In:
INTERNATIONAL CONGRESS ON ANIMAL REPRODUCTION AND ARTIFICIAL INSEMINATION, 11. Proceedings... Dublin, v.5,
p.117-125, 1988.
BRUSTOLINI, P.C. et al. Efeitos de diferentes fontes lipídicas e níveis de
energia sobre o desempenho reprodutivo de marrãs. Arq. Bras. Med. Vet. Zootec. v. 56 (4), 2004.
BIRD, I.M.; ZHANG, L.B.; MAGNESS, R.R. Possible mechanisms underlying
pregnancy-induced changes in uterine artery endothelial function, Am. J. Physiol.,
v. 284, pp. R245-R258, 2003.
CAMERON, J.L. Nutritional determinants of puberty. Nut. Rev., v. 54, p. S17-S22,
1996.
CARTER, S.D.; HILL, G.M.; MAHAN, D.C.; NELSSEN, J.L.; RICHERT, B.T.;
SHURSON, G.C. Effects of dietary valine concentration
on lactational performance of sows nursing large
litters. J. Anim. Sci., v. 78, p.2879-2884, 2000.
CLOSE, W.H.; COLE, D.J.A.
The pre-breeding gilt. In: Nutritional of Sows and Boars.
CLOSE, W.H.; COLE, D.J.A. Nutrition of sows and boars. 1rst.
Ed.Nottinghan:
CLOWES, E.J.; AHERNE, F.X.; FOXCROFT, G.R.; BARACOS, V.E. Selective
protein loss in lactating sows is associated with reduced litter growth and
ovarian function. J. Anim. Sci.,
v.81, p. 753-764, 2003b.
COSGROVE. J.R. et al.
Gonadotrophic-independent mechanisms participate in
ovarian responses to realimentation in
feed-restricted prepubertal gilts. Biol. Reprod.,
v. 47, p. 739, 1992.
DAVIS, T.A.; NGUYEN, H.V.; GARCIAA-BRAVO, R.; FIOROTTO, M.L.; JACKSON,
E.M.; LEWIS, D.S.; LEE, D.R.; REEDS, P.J. Amino acid composition of human milk
is not unique, J. Nutr.,
v. 124, pp. 1126-1132, 1994.
DRITZ, S. Replacement gilt nutrition. In: AMERICAN ASSOCIATION OF SWINE
VETERINARIANS 37th ANNUAL MEETING 2006. Proceedings... p29-35, 2006.
DWYER, C.M.,
ESCOBAR, J. Modeling changes in the body composition of primiparous lactating sows as affected by four dietary
regimens. M.S. thesis.
FOXCROFT, G.R. Interactions between body composition,
metabolic state and reproduction in swine. In: WESTERN
NUTRITION CONFERENCE, 11. Proceedings...
1990.
FOXCROFT, G.R. Female reproduction. In:
FOXCROFT, G.R. (Ed). Reproduction
in domesticated animals.
FOXCROFT, G.R.;COSGROVE, J.R.; AHERNE, F.X.
Relationship between metabolism and reproduction. In: INTERNATIONAL PIG VETERINARY SCIENCE CONGRESS, 14,
1996,
FOXCROFT, G.R. Recognizing the characteristics
of our new dam lines. Proceedings of Allen D. Leman Swine
Conference,
FRANK, J.W.; ESCOBAR, J.; NGUYEN, H.V.; JOBGEN, S.C.; JOBGEN, W.S.;
DAVIS, T.A.; WU., G. OralN-carbamylglutamate
supplementation increases protein synthesis in skeletal muscle of piglets. J. Nutr., v.137, p.315-319, 2007.
GAINES, A.M.; BOYD, R.D.; JOHNSTON, M.E.; USRY, J.L.; TOUCHETTE, K.J.;
ALLEE, G.L. Effects of dietary valine concentration
on lactational performance of sows nursing large
litters. NCR-42 Committe on Swine
Nutrition. J. Anim.
Sci., v.78, p.2879-2884, 2000.
HEAD, R.H. & WILLIAMS, I.H. Mammogenesis
is influenced by pregnancy nutrition. In: III MANIPULATING PIG PRODUCTION, Batterham, E.S. Ed. Australasian
Pig Science
Associatinon, Qttword, p.76, 1994.
HOSTETLER, C.E.,. KINCAID,
R.L.; MIRANDO, M.A. The role of essential trace
elements in embryonic and fetal development in livestock. Vet. J., v.166, p.125-139. 2003.
HUGHES, P.E. The effects of food level during lactation and early
gestation on the reproductive performance of mature sows. Anim. Prod. V. 57, p. 437-445, 1993.
JONES, D.B.; STAHLY, T.S. Impact of amino acid
nutrition during lactation on subsequent reproductive function of sows. J. Anim. Sci.,
v. 73 (Suppl. 1), p.183 (Abstr.),
1995.
JONES, D.B.; STAHLY, T.S. Impact of amino acid
nutrition during lactation on body nutrient mobilization and milk nutrient
output in primiparous sows. J. Anim. Sci.,
v. 77, p.1513-1522, 1999.
KEMP. B. et al. Effects of ennergy source in the diet on reproductive hormones and
insulin during lactation and subsequent estrus in multiparous
sows. J. Anim. Sci., v. 73, p.
3022-3029, 1995.
Kim, S.W.; Wu, G. Amino acid requirements for breeding
Sows. In: II Simpósio Internacional sobre
exigências nutricionais de aves e suínos - UFV, p.199-218, 2005.
Kim, S. W.; Easter, R.A. Nutrient mobilization from
body tissues as influenced by litter size in lactating sows. J.
Anim. Sci., v.78,
p.2172-2178, 2001.
KIM, S.W., HURLEY, W.L.; HAN, I.K.; EASTER, R.A. Growth
of nursing pigs related to the characteristics of nursed mammary glands. J. Anim. Sci.,
v. 78, p.1313-1318. 2000.
KIM, S. W.; EASTER, R.A. Nutrient mobilization from
body tissues as influenced by litter size in lactating sows. J. Anim. Sci.,
v. 78, p.2172-2178, 2001.
KIM, S. W.; EASTER, R.A. Amino acid utilization for reproduction in sowsIn: Amino Acids in Animal Nutrition. 2nd
ed. J. P. F. D Mello, ed. CAB International,
KIM, S.W.; MCPHERSON, R.L.; WU, G. Dietary arginine
supplementation enhances the growth of milk-fed young pigs, J. Nutr., v.
134, pp. 625-630, 2004.
KIM, S W.; WU, G. Dietary arginine
supplementation enhances the growth of milk fed young pigs. J. Nutr.,
v.134, p.625-630, 2004.
KIM, S.W.; MATEO, R.D.; YIN, Y.L.; WU, G. Functional amino acids and
fatty acids for enhancing production performance of sows and piglets, Asian-Aust. J. Anim. Sci., v. 20,
pp. 295-306, 2007.
KING, R.H.; TONER, M.S.; DOVE, H.; ATWOOD, C.S.;
BROWN. W.G. The response
of first-litter sows to dietary protein level during lactation. J Anim
Sci., v. 71, p. 2457-2463, 1993.
KITT, S.J.; MILLER, P.S.; FISCHER, R.L. Effects of sow
dietary glutamine supplementation on sow and litter performance, subsequent
weanling pig performance and intestinal development after an immune challenge. Nebraska Swine Report, p.14-17,
2004.
KNOPF, R.F.; CONN, J.W.; FAJANS, S.S.; FLOYD, J.C.; BUNTSCHE, E.W.;
REILL, J.A. Plasma growth hormone response to intravenous administration of
amino acids. J. Clin. Endocrinol. Metab.,
v. 25, p.1140-1144, 1968.
LACASSE, P.; PROSSER, C.G. Mammary blood flow does not limit milk yield
in lactating goats. J. Dairy Sci., v.86, p.2094-2097,
2003.
LI, P.; YIN, Y.L.; LI, D.F.; KIM, S.W.; WU, G. Amino acids and immune
function, Br. J. Nutr.,
v.98, pp. 237-252, 2007MAHAN, D.C., FETTER, A.W. Dietary calcium and phosphorus
levels for reproducing sows. J. Anim. Sci., v. 34, p.283,1982.
LOBLEY, G.E.; HOSKIN, S.O.; McNEIL,
C.J. Glutamine in Animal Science and Production. Journal
of Animal Science, v.131, p.525S-2531S, 2001.
MACHADO, G.S. Efeitos de diferentes
fontes de energia sobre a eficiência reprodutiva de marrãs cíclicas.
2005. 151 p. Tese (Doutorado
MAHAN, D. The
Changing Mineral Status of High Producing Sows - What are their needs and when
are the critical periods? In: Swine Nutrition Conference
Proceeding. Indianápolis, Indiana, p. 17-27, 2006.
MANSO,H.C.C.C. Avaliação da glutamina sintetase e da concentração a
glutamina no terço final da gestação e na lactação de camundongos fêmeas e
matrizes suínas primíparas. Tese de doutorado - Universidade Federal Rural de
Pernambuco, 2006.
MATAMOROS, I.A. et al. Effects of exogenous insulin and
body condition on metabolic hormones and gonadotropin-induced
follicular development in prepurtal gilts. J. Anim. Sci., v.69, p.2081-2091, 1991.
MATEO, R.D.; WU, G.; BAZER, F.W.; PARK, J.C.; SHINZATO, I.; KIM, S.W.
Dietary l-arginine supplementation enhances the
reproductive performance of gilts, J. Nutr., v. 137, p. 652-656, 2007.
MATEO, R.D.; WU, G.; MOON, H.K.; CARROLL, J.A.; KIM, S.W. Effects of
dietary arginine supplementation during gestation and
lactation on the performance of lactating primiparous
sows and nursing piglets. J
Anim Sci., v. 86,
p.827-835, 2008.
MAXSON, P.F.,
McDONALD, L.E. Veterinary
endocrinology and reproduction. 3
ed., Filadélfia: Lea & Febiger,
p. 560, 1980.
MCPHERSON, R.L.; JI, F.; WU, G.; KIM, S.W. Fetal growth and
compositional changes of fetal tissues in the pigs, J. Anim. Sci., v.
82, pp. 2534-2540, 2004.
MEININGER, C.J.; WU, G. Regulation of endothelial cell
proliferation by nitric oxide. Methods
Enzymol., v. 352, p.280-295, 2002.
MURGAS, L.D.S. Efeito do consumo de
energia na fase pré-puberal sobre os desempenhos produtivo e reprodutivo de
marrãs. 1994. 65p. Dissertação (Mestrado) - Universidade Federal de
Viçosa, Viçosa.
NEWSHOLME, P.; BRENNNAN, L.; RUBI, B.; MAECHLER, P.New insights into amino
acid metabolism, beta-cell function and diabetes, Clin. Sci., v. 108, p. 185-194, 2005.
NIMMO. R.D., PEO, E. R.; JR.,
MOSER, B D.; LEWIS, A. J. Effect of level of dietary calcium - phosphorus
during growth and gestation on performance, blood and bone parameters of swine.
J. Anim. Sci.,
v. 52, p.1330, 1981.
NOBLET, J.; CLOSE, W.H.; HEAVENS, R.P.; BROWN, D.
Studies on the energy metabolism of the pregnant sow. 1. Uterus and mammary tissue development. Br. J. Nutr. , v. 53, p.251-265, 1985.
NOBLET, J.; DOURMAD, J.Y.; ETIENNE, M. Energy utilization in pregnant
and lactating sows: modeling of energy requirements. J Anim Sci., v.68, p.562-572, 1990.
NOBLET, J. & ETIENNE, M. Energetic efficiency of milk production. In:
VERSTEGEN, M.W.A.; MOUGHAN,P,J. & SCHRAMA, J.W. The Lactating Sow. Wagening Pers,
NRC. Nutrient
Requirements of Domestic Animals. Nutrient Requirements of Swine.8th
Revised
NRC. Nutrient
Requirements of Swine. 9th ed. National Academy Press,
NRC. Nutrient
Requirements of Domestic Animals. Nutrient Requirements of Swine.10th
Revised
PETERS, J. C. Evaluating the efficacy of dietary organic and inorganic
trace minerals in reproducing female pigs on reproductive performance and body
mineral composition. PhD thesis, 2006.
PIC
PORETSKY, L., KALIN, M.F. The gonadotropic
function of insulin. Endocrine
reviews, v.8, p. 132-141, 1987
RAMAEKERS, P.; KEMP, B.; VAN DER LENDE, T.Progenos
in sows increases number of piglets born, J.
Anim. Sci., v. 84
(Suppl. 1), p. 394., 2006.
REVELL, D.K.; WILLIAMS, I.H.; MULLAN, B.P.; RANDFORD, J.L.; SMITS, R.J.
Body composition at farrowing and nutrition during
lactation affect the performance of primiparous sows:
I. Voluntary feed intake, weight loss, and plasma metabolites J. Anim. Sci., v.76, p.1729- 1737,
1998.
REYNOLDS, L.P.; REDMER, D.A Angiogenesis in the
placenta, Biol. Reprod.,
v. 64, pp. 1033-1040, 2001.
Richert, B.T., Goodband, R.D.; Tokach, M.D.; Nelssen, J.L. Increasing valine, isoleucine, and total branched-chain amino acids for lactating
sows. J. Anim. Sci., v.75,
p.2117-2128, 1997a.
Richert, B.T., Tokach, M.D.; Goodband, R.D.; Nelssen, J.L.; Campbell, R.G.; Kershaw, S. The effect of
lysine and valine fed during lactation on sow and
litter performance. J. Anim. Sci.,
v.75, p.1853-1860, 1997b.
Richert, B.T.; Tokach, M.D.; Goodband, R.D.; Nelssen, J.L.; Pettigrew, J.E.; Walker, R.D.; Johnston,
L.J. Valine requirement of the high-producing
lactating sow. J. Anim. Sci.,
v. 74, p.1307-1313, 1996.
RICHERT, B.T.; TOKACH, M.D.; GOODBAND, R.D.; NELSSEN, J.L.; PETTIGREW,
J.E.; WALKER, R.D.; JOHNSTON,L.J. Valine requirement
of the high-producing lactating sow. J Anim Sci., v.74,
p.1307-1313, 1996.
ROSTAGNO, H.S., ALBINO, L.F.T., DONZELE, J.L. et al. Tabelas
brasileiras para aves e suínos. Composição de alimentos e exigências
nutricionais. Viçosa: UFV. 141p. 2000.
ROTH, F.X.; FICKLER, J.; KIRCHGESSNER, M. Effect of dietary arginine and
glutamic acid supply on the N-balance of piglets. 5.
Communication on the importance of nonessential amino acids for protein
retention, J. Anim.
Physiol. Anim. Nutr., v. 73, p. 202-212, 1995.
SCHWARTZ, M.W. et al. Insulin in the brain: a hormonal regulator of
energy balance. Endoc. Rev., v.13, p.
387-414, 1992.
SOBESTIANSKY, J.; BARCELLOS, D.E.S.N.; SESTI, L.A.C. Introdução de animais
em um sistema de produção. In: SOBESTIANSKY, J.; WENTZ, IVO; SILVEIRA, P.R.S.;
SESTI, L.A. (Eds). Suinocultura itensiva. Comcórdia-SC:
Embrapa-CNPSA.1998. cap.19, p. 337-348.
SOUTHERN, L.L.; BAKER, D.H. Arginine
requirement of the young pig, J. Anim. Sci., v.57, p.
402-412, 1983.
URSCHEL, K.L.; SHOVELLER, A.K.; UWIERA, R.R.; PENCHARZ, P.B.; BALL, R.O.
Citrulline is an effective arginine
precursor in enterally fed neonatal piglets, J. Nutr.,
v.136, p.1806-1813, 2006.
VAN DEN BRAND, H. et al. Effects of postweaning
dietary energy source on reproductive traits in primiparous
sows. J.Anim. Sci.,
v. 79, p. 420-426, 2001.
WELDON, W.C., THUILIN, A.J., Mac DOUGALD, O.A., et al..
Effects of increased dietary energy and protein during late
gestation on mammary development in gilts. J. Anim. Sci. ,
69:194-200, 1991.
WHITEMORE, C. T. Review - Nutrition reproduction interactions in primiparous sows. Livest. Product. Sci., v. 46, p. 65-83, 1996.
WU, G.; KNABE, D.A. Free and protein-bound amino acids
in sow's colostrum and milk. J.
Nutr., v.124, p.415-424, 1994.
WU, G. Effects of concanavalin
A and phorbol myristate
acetate on glutamine metabolism and proliferation of porcine intraepithelial
lymphocytes, Comp. Biochem. Physiol., v.114A, pp. 363-368, 1996.
WU, G.; MORRIS JR., S.M. Arginine metabolism:
nitric oxide and beyond, Biochem. J. 336, pp. 1-17, 1998.
WU, G; OTT, T.L.; KNABE, D.A.; BAZER, F.W. Amino acid composition of the
fetal pig, J. Nutr.,
v.129, p. 1031-1038, 1999.
WU, G.; MEININGER, C.J. Arginine
nutrition and cardiovascular function, J. Nutr., v. 130, pp. 2626-2629, 2000.
WU, G.; BAZER, F.W.; CUDD, T.A.; MEININGER, C.J.; SPENCER, T.E. Maternal
nutrition and fetal development, J. Nutr., v. 134 , p. 2169-2172, 2004.
WU, G. Nutrição da glutamina em suínos: da pesquisa básica à suinocultura. Disponível em: <www.lisina.com.br>. Acesso em: 26
de dez. 2005.
WU, G.; BAZER, F.W.; WALLACE, J.M.; SPENCER, T.E. Intrauterine growth
retardation: implications for the animal sciences, J. Anim. Sci., v.
84, pp. 2316-2337, 2006.
ENGORPART POR 20100206 ENGORPART POR 20100206